



Introduction
In the process of aging, human brain displays a gradually enhanced inflammatory status called ‘inflamm-aging’ [1]. Although this status is crucial for the brain to remove senescent cells and extrinsic pathogenic substances, recent evidence has proven that chronic neuroinflammation is detrimental for the brain and may contribute to the etiology of various aging-related neurodegenerative diseases such as Alzheimer’s disease (AD) and Parkinson’s disease (PD) [2-4]. This notion was supported by previous findings that anti-inflammatory therapies could halt the progression of AD and PD in animal models [5,6].
Microglia are the major type of immune cells in the brain [7]. During the aging process, microglia can be activated and contribute to the neuroinflammation via release of pro-inflammatory cytokines [8,9]. This inflammatory response may subsequently lead to the injury or even death of neurons and synapses [10,11]. Therefore, microglia may represent a principal target for restricting the excessive neuroinflammation under disease status.
The renin-angiotensin system (RAS) has been accepted as an important modulator in the circulation system for maintaining the homeostasis of sodium and water [12]. In addition to the circulation system, cumulative evidence indicated that RAS also exists in colon, lungs, liver and brain, and exerts important physiological functions in these organs and tissues [13,14]. As a newly identified heptapeptide of RAS, angiotensin-(1-7) (Ang-(1-7)), along with angiotensin converting enzyme (ACE) 2 and MAS1 receptor, constitutes ACE2/Ang-(1-7)/MAS1 axis [15]. Emerging evidence revealed a crucial role of this axis in regulating inflammatory responses under various pathological conditions such as colitis, lung fibrosis, hepatic steatosis and ischemic stroke [16-19]. However, the relationship of brain ACE2/Ang-(1-7)/MAS1 axis with aging-related neuroinflammation is less studied thus far.
In this study, by using senescence-accelerated mouse prone 8 (SAMP8) mice, an animal model of accelerated aging [20,21], we revealed that the neuroinflammation in the aged brain might be attributed to a decreased level of Ang-(1-7). More importantly, we provided evidence that AVE0991, a nonpeptide analogue of Ang-(1-7), attenuated the aging-related neuroinflammation via suppression of microglial-mediated inflammatory response through a MAS1 receptor-dependent manner. Meanwhile, this protective effect might be ascribed to the M2 activation of microglia induced by AVE0991. Taken together, these findings reveal the association of Ang-(1-7) with the inflammatory response in the aged brain and uncover the potential of its nonpeptide analogue AVE0991 in attenuation of aging-related neuroinflammation.
Results
Inflammatory markers are increased in the brains of SAMP8 mice during the aging process
First, the dynamic changes of inflammatory markers including IL-1β, IL-6, and TNF-α in the brains of 4-, 8-, and 12-month-old SAMP8 mice as well as their age-matched controls were investigated by qRT-PCR. As shown in Figure 1, the mRNA levels of Il1b, Il6 and Tnf were significantly increased in the brains of SAMP8 mice during the aging process (n=8, P<0.05). Meanwhile, a significant higher expression of Il1b, Il6 and Tnf was observed in the brains of 8- and 12-month-old SAMP8 mice when compared with their age-matched controls (Figure 1, n=8, P<0.05). These findings indicated an increased neuroinflammatory response in the aged brain.

Figure 1. Inflammatory markers are increased in the brains of SAMP8 mice during the aging process. (A) The dynamic changes of Il1b mRNA levels in the brains of 4-, 8-, and 12-month-old SAMP8 mice as well as their age-matched SAMR1 control mice were investigated by qRT-PCR. (B) The dynamic changes of Il6 mRNA levels in the brains of 4-, 8-, and 12-month-old SAMP8 mice as well as their age-matched SAMR1 control mice were investigated by qRT-PCR. (C) The dynamic changes of Tnf mRNA levels in the brains of 4-, 8-, and 12-month-old SAMP8 mice as well as their age-matched SAMR1 control mice were investigated by qRT-PCR. Gapdh was used as an internal control. All data were analyzed by one-way ANOVA followed by Tukey’s post hoc test and were expressed as a fold change relative to 4-month-old SAMR1 control mice. Columns represent mean ± SD (n=8 per group). *P<0.05. #P<0.05 versus age-matched SAMR1 control mice.
Ang-(1-7) levels are decreased in the brains of SAMP8 mice during the aging process
Next, we determined whether this increased neuroinflammatory response was related to any alterations in the components of brain ACE2/Ang-(1-7)/MAS1 axis. As shown in Figure 2, in the brains of SAMP8 mice, the levels of Ang-(1-7) were markedly decreased during the aging process (n=8, P<0.05). Meanwhile, the Ang-(1-7) levels in the brains of 8- and 12-month-old SAMP8 mice were significantly lower when compared with those of their age-matched controls (n=8, P<0.05). It should be noted that the activity of ACE2 and the mRNA level of Mas1 remained stable in the brains of SAMP8 mice during the aging process. These findings implied that the increased neuroinflammatory response might be attributed to the decreased Ang-(1-7) levels in the aged brain.
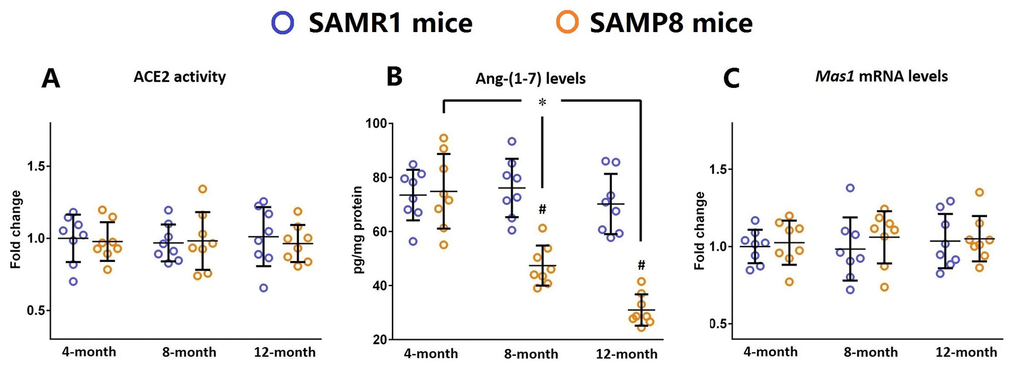
Figure 2. Ang-(1-7) levels are decreased in the brains of SAMP8 mice during the aging process. (A) The activity of ACE2 in the brains of 4-, 8-, and 12-month-old SAMP8 mice as well as their age-matched SAMR1 control mice were assessed using a specific detection kit. (B) The Ang-(1-7) levels in the brains of 4-, 8-, and 12-month-old SAMP8 mice as well as their age-matched SAMR1 control mice were detected by ELISA. (C) The expression of Mas1 mRNA levels in the brains of 4-, 8-, and 12-month-old SAMP8 mice as well as their age-matched SAMR1 control mice were assessed by qRT-PCR. Gapdh was used as an internal control. Data from panel B and C were expressed as a fold change relative to 4-month-old SAMR1 control mice. All data were analyzed by one-way ANOVA followed by Tukey’s post hoc test. Columns represent mean ± SD (n=8 per group). *P<0.05. #P<0.05 versus age-matched SAMR1 control mice.
AVE0991 attenuates neuroinflammation in the brains of SAMP8 mice
Based on the above findings, we then hypothesized that restoration of Ang-(1-7) levels might attenuate neuroinflammation in the aged brain. Since Ang-(1-7) will be rapidly inactivated and degraded by several proteases and thus has a relatively short duration of biological effect in vivo, we employed AVE0991, a nonpeptide analogue of Ang-(1-7) [22], in the subsequent experiments.
As illustrated by Supplementary Figure S1, eight-month-old SAMP8 mice were injected intraperitoneally with vehicle or AVE0991 (1, 3 or 10 mg/kg/day) once a day for 30 consecutive days. AVE0991 injection (3 or 10 mg/kg/day) significantly decreased the mRNA levels of Il1b, Il6 and Tnf in the brains of SAMP8 mice (Figure 3A-C, n=8, P<0.05). In addition, a corresponding decrease in protein levels of IL-1β, IL-6 and TNF-α in the brains of SAMP8 mice was observed following AVE0991 (3 or 10 mg/kg/day) injection (Figure 3D-F, n=8, P<0.05). It should be noted that AVE0991 (1, 3 or 10 mg/kg/day) had no significant effect on mRNA levels of Il1b, Il6 and Tnf in the brains of 8-month-old SAMR1 control mice (Preliminary experiments, Supplementary Figure S2, n=5). These findings indicated that AVE0991 attenuated neuroinflammation in the aged brain.
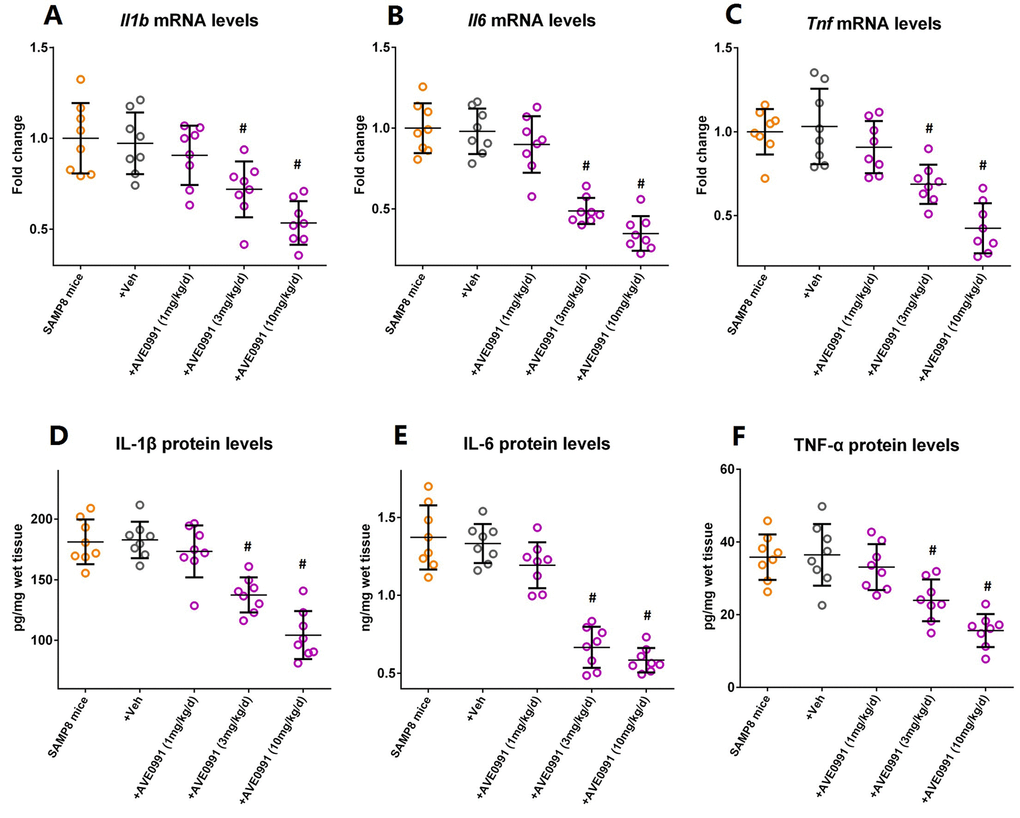
Figure 3. AVE0991 attenuates neuroinflammation in the brains of SAMP8 mice. Eight-month-old SAMP8 mice were injected intraperitoneally with vehicle or AVE0991 (1, 3 or 10 mg/kg/day) for 30 days. Afterwards, mice were sacrificed for analysis. (A) The mRNA levels of Il1b in the brains were investigated by qRT-PCR. (B) The mRNA levels of Il6 in the brains were investigated by qRT-PCR. (C) The mRNA levels of Tnf in the brains were investigated by qRT-PCR. (D) The protein levels of IL-1β in the brains were investigated by ELISA. (E) The protein levels of IL-6 in the brains were investigated by ELISA. (F) The protein levels of TNF-α in the brains were investigated by ELISA. In panel A-C,Gapdh was used as an internal control, and data were expressed as a fold change relative to non-treated SAMP8 mice. All data were analyzed by one-way ANOVA followed by Tukey’s post hoc test. Columns represent mean ± SD (n=8 per group). #P<0.05 versus vehicle-treated SAMP8 mice.
AVE0991 suppresses microglial-mediated inflammatory response through a MAS1 receptor-dependent manner
Afterwards, we tried to elucidate the underlying mechanisms by which AVE0991 attenuated neuroinflammation. As MAS1 receptor was reported to be expressed by microglia whilst activation of microglia contributed to the neuroinflammatory response during the aging process [8,23], we then speculated that AVE0991 attenuated neuroinflammation by suppressing microglial-mediated inflammatory response. To test this hypothesis, primary microglia were isolated from the brain of 8-month-old SAMP8 mice. Double immunofluorescence staining indicated a good co-localization of MAS1 with a microglial marker AIF1, confirming that MAS1 receptor is expressed by microglia (Figure 4A). Next, microglia were treated for 4 h with 100 ng/ml LPS with or without 4 h pre-incubation with AVE0991 (1×10-8, 1×10-7 or 1×10-6 M) in the presence or absence of A-779 (1×10-6 M). LPS incubation induced an inflammatory response in microglia, since the mRNA levels of Il1b, Il6 and Tnf were significantly increased (Figure 4B-D, P<0.05). Pre-incubation with AVE0991 (1×10-7 or 1×10-6 M) attenuated the LPS-induced increase in Il1b and Tnf mRNA levels (Figure 4B-D, P<0.05). Meanwhile, the increase in Il6 mRNA levels was markedly inhibited by AVE0991 (1×10-8, 1×10-7 or 1×10-6 M). It is worthy to note that the effects of AVE0991 (1×10-7 M) on the mRNA levels of Il1b, Il6 and Tnf were fully abolished by A-779 (1×10-6 M), an antagonist of MAS1 receptor (Figure 4B-D, P<0.05). In addition, A-779 (1×10-6 M) alone did not significantly influence the expression of these inflammatory markers (Figure 4B-D).
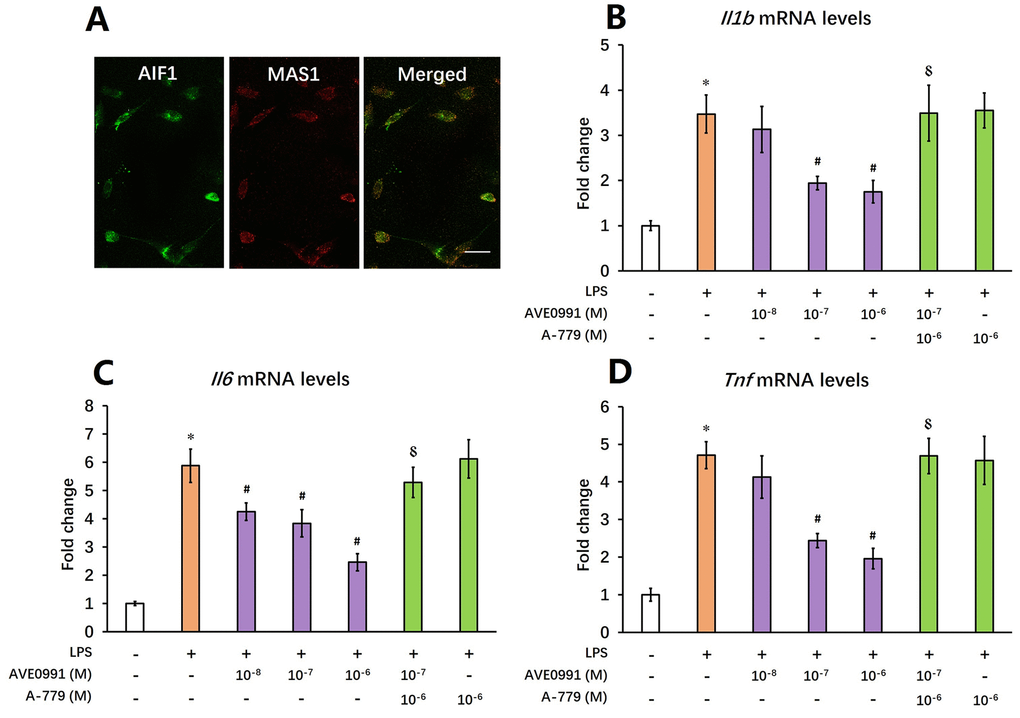
Figure 4. AVE0991 suppresses microglial-mediated inflammatory response through a MAS1 receptor-dependent manner. Primary microglia were directly isolated from the brain of 8-month-old SAMP8 mice. (A) The localization of MAS1 receptor on microglia was confirmed by double immunofluorescence staining. Scale bar=15 μm. Next, microglia were treated for 4 h with 100 ng/ml LPS with or without 4 h pre-incubation with AVE0991 (1×10-8, 1×10-7 or 1×10-6 M) in the presence or absence of A-779 (1×10-6 M) and were harvested and lysed for analysis. (B) The mRNA levels of Il6 were investigated by qRT-PCR. (C) The mRNA levels of Ilb were investigated by qRT-PCR. (D) The mRNA levels of Tnf were investigated by qRT-PCR. In panel B-D,Gapdh was used as an internal control, and data were expressed as a fold change relative to non-treated microglia. All data were analyzed by one-way ANOVA followed by Tukey’s post hoc test. Columns represent mean ± SD (n=4-6). *P<0.05 versus non-treated microglia. #P<0.05 versus microglia incubated with LPS alone. ⸹P<0.05 versus microglia treated with LPS+AVE0991 (1×10-7M).
AVE0991 promotes M2 activation of microglia
As reduced inflammatory response represents a major characteristic of M2 activation of microglia [24], we then hypothesized that AVE0991 suppressed microglial-mediated inflammatory response by switching microglia toward the M2 activation. To test this hypothesis, we measured the mRNA levels of M2 activation makers including Arg1, Il10, and Retnla in the brains of SAMP8 mice following AVE0991 treatment. Injection of AVE0991 (1, 3 or 10 mg/kg/day for 30 consecutive days) markedly increased the mRNA levels of Arg1 in the brains of SAMP8 mice (Figure 5A, P<0.05). Meanwhile, the mRNA levels of Il10 and Retnla in the brains of SAMP8 mice were significantly elevated following AVE0991 injection (3 or 10 mg/kg/day for 30 consecutive days, Figure 5B and C, P<0.05). To further validate this finding, microglia isolated from 8-month-old SAMP8 mice were treated for 4 h with 100 ng/ml LPS with or without 4 h pre-incubation with AVE0991 (1×10-8, 1×10-7 or 1×10-6 M) in the presence or absence of A-779 (1×10-6 M). As shown in Figure 6A and C, pre-incubation with AVE0991 (1×10-7 or 1×10-6 M) significantly increased the mRNA levels of Arg1 and Retnla (P<0.05). Meanwhile, the mRNA levels of Il10 were markedly elevated by AVE0991 (1×10-8, 1×10-7 or 1×10-6 M, Figure 6B, P<0.05). It should be noted that the effects of AVE0991 (1×10-7 M) on Arg1, Il10, and Retnla mRNA levels were fully reversed by A-779 (1×10-6 M, Figure 6, <0.05). Additionally, A-779 (1×10-6 M) itself did not markedly affect the expression of these M2 activation markers (Figure 6).
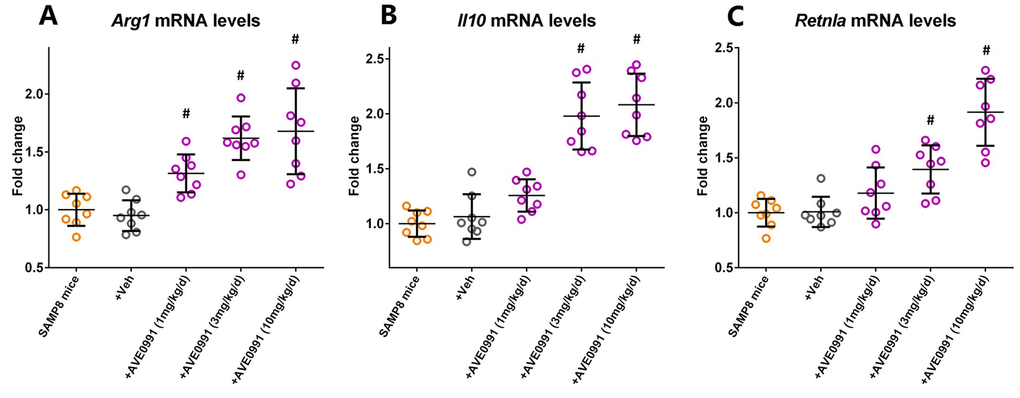
Figure 5. AVE0991 elevates microglial M2 activation makers in the brains of SAMP8 mice. Eight-month-old SAMP8 mice were injected intraperitoneally with vehicle or AVE0991 (1, 3 or 10 mg/kg/day) for 30 days. Afterwards, mice were sacrificed for analysis. (A) The mRNA levels of Arg1 in the brains were investigated by qRT-PCR. (B) The mRNA levels of Il10 in the brains were investigated by qRT-PCR. (C) The mRNA levels of Retnla in the brains were investigated by qRT-PCR. Gapdh was used as an internal control, and data were expressed as a fold change relative to non-treated SAMP8 mice. All data were analyzed by one-way ANOVA followed by Tukey’s post hoc test. Columns represent mean ± SD (n=8 per group). #P<0.05 versus vehicle-treated SAMP8 mice.
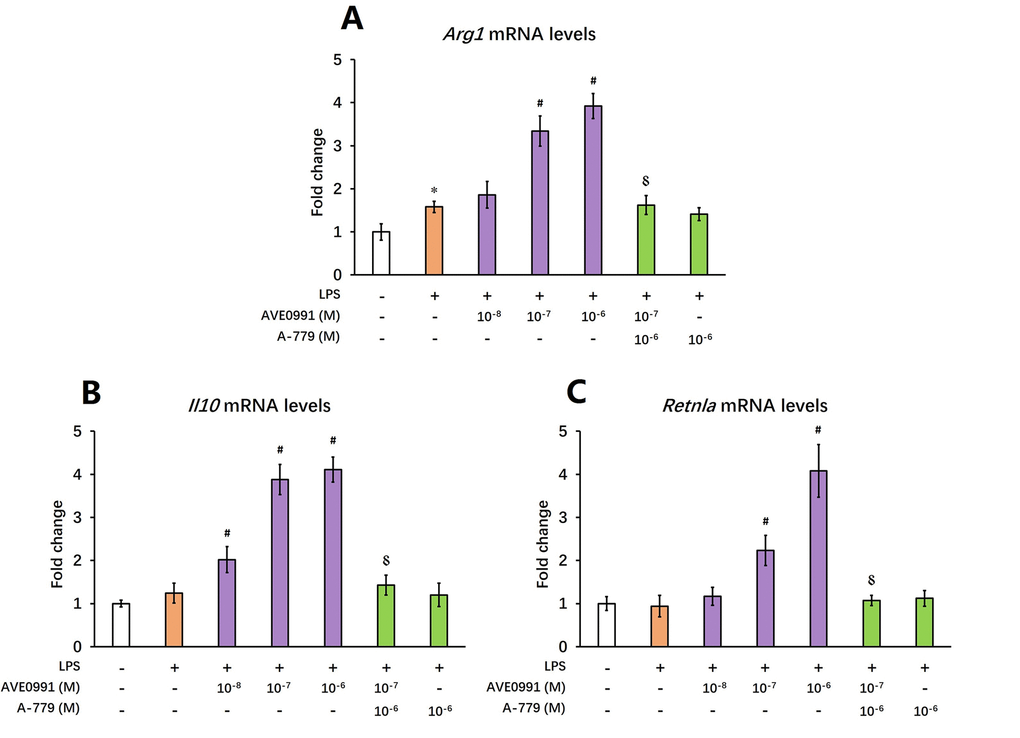
Figure 6. AVE0991 elevates M2 activation makers in primary microglia. Primary microglia were directly isolated from the brain of 8-month-old SAMP8 mice. They were treated for 4 h with 100 ng/ml LPS with or without 4 h pre-incubation with AVE0991 (1×10-8, 1×10-7 or 1×10-6 M) in the presence or absence of A-779 (1×10-6 M) and were harvested and lysed for analysis. (A) The mRNA levels of Arg1 were investigated by qRT-PCR. (B) The mRNA levels of Il10 were investigated by qRT-PCR. (C) The mRNA levels of Retnla were investigated by qRT-PCR. Gapdh was used as an internal control, and data were expressed as a fold change relative to non-treated microglia. All data were analyzed by one-way ANOVA followed by Tukey’s post hoc test. Columns represent mean ± SD (n=4-6). *P<0.05 versus non-treated microglia. #P<0.05 versus microglia incubated with LPS alone. ⸹P<0.05 versus microglia treated with LPS+AVE0991 (1×10-7M).
Discussion
First, in this study, we showed a progressively enhanced inflammatory response in the aged brain, since the expression of inflammatory markers including IL-1β, IL-6, and TNF-α was significantly increased in SAMP8 mice brain during the aging process. These results were compatible with previous findings from our group and others that SAMP8 mice exhibited a chronic neuroinflammatory status [25,26]. Meanwhile, we demonstrated that the levels of Ang-(1-7) was significantly reduced in the aged brain, and these findings were in accordance with our previous observations in SAMP8 mice [27].
As a recently identified bioactive peptide of RAS, Ang-(1-7) is produced mainly from angiotensin II by ACE2 and binds to MAS1 receptor for its signaling and biologic functions [28]. Emerging evidence suggested that Ang-(1-7) could inhibit inflammatory responses under various pathological conditions. In an experimental model of colitis, daily Ang-(1-7) treatment significantly ameliorated inflammation induced by dextran sulfate sodium [16]. Meanwhile, the anti-inflammatory effects of Ang-(1-7) were also observed in a mouse model of lung fibrosis induced by bleomycin [17]. Moreover, treatment with an oral formulation of Ang-(1-7) suppressed the expression of inflammatory markers and improved hepatic functions in a mouse model of hepatic steatosis caused by a high-fat diet [18]. In addition, in a rat model of ischemic stroke, Ang-(1-7) infusion exerted neuroprotective effects via inhibition of NF-κB-mediated inflammatory response [19]. In view of the anti-inflammatory actions of Ang-(1-7), we speculated that the enhanced inflammatory response in the brain during the aging process might attributed to the reduction of Ang-(1-7) levels, and restoration of Ang-(1-7) levels might attenuate this aging-related neuroinflammation.
However, it should be noted that Ang-(1-7) has a relatively short duration of biological effect in vivo since it can be rapidly inactivated and degraded by several proteases such as ACE and neutral endopeptidase [29-31]. To avoid theses disadvantages of Ang-(1-7), we then employed its nonpeptide analogue AVE0991 in the subsequent experiments to further verify our hypothesis [22]. As expected, AVE0991 treatment ameliorated the aging-related neuroinflammation in SAMP8 mice, since the expression of inflammatory markers was markedly reduced. To our knowledge, this is the first study to reveal the anti-inflammatory property of AVE0991 in an animal model of accelerated aging.
Afterwards, we tried to elucidate the underlying mechanisms by which AVE0991 attenuated neuroinflammation. As revealed by our double immunofluorescence staining, MAS1 receptor was expressed by microglia, and this observation was supported by an earlier study from Liu and colleagues showing that MAS1 receptor was stably expressed by human microglia [23]. As the main immune cells in the brain, microglia was activated during the aging process and thus contributed to the chronic neuroinflammation via release of pro-inflammatory cytokines [8,9]. In this study, we revealed that AVE0991 treatment suppressed the LPS-induced expression of inflammatory markers in primary microglia isolated from SAMP8 mice, and this effect can be completely abolished by an antagonist of MAS1 receptor A-779. This observation indicated that AVE0991 attenuated neuroinflammation by suppressing microglial-mediated inflammatory response via a MAS1 receptor-dependent manner. More interestingly, an increased level of microglial M2 activation markers including Arg1, Retnla and Il10 was observed following AVE0991 treatment. In contrast to M1 activation, M2 activation of microglia is featured by a suppressed inflammatory response as well as an enhanced phagocytic activity [24]. Therefore, it seemed that the anti-inflammatory actions of AVE0991 were achieved by switching microglia toward the M2 activation in this scenario.
It is noteworthy that the current study also has some limitations. For example, SAMP8 mouse used in this study is an animal model of accelerated aging. Its pathophysiological process and neuropathology are not equal to those under normal aging conditions [20,21]. Therefore, the current findings in this study need to be further confirmed using normal aging mice in future.
Summarily, in this study, we revealed that the neuroinflammation in the aged brain might be attributed to a decreased level of Ang-(1-7). More importantly, we provided evidence that AVE0991, a nonpeptide analogue of Ang-(1-7), attenuated the aging-related neuroinflammation via suppression of microglial-mediated inflammatory response through a MAS1 receptor-dependent manner. Meanwhile, this protective effect might be ascribed to the M2 activation of microglia induced by AVE0991 (Summarized in Figure 7). Taken together, these findings reveal the association of Ang-(1-7) with the inflammatory response in the aged brain and uncover the potential of its nonpeptide analogue AVE0991 in attenuation of aging-related neuroinflammation.
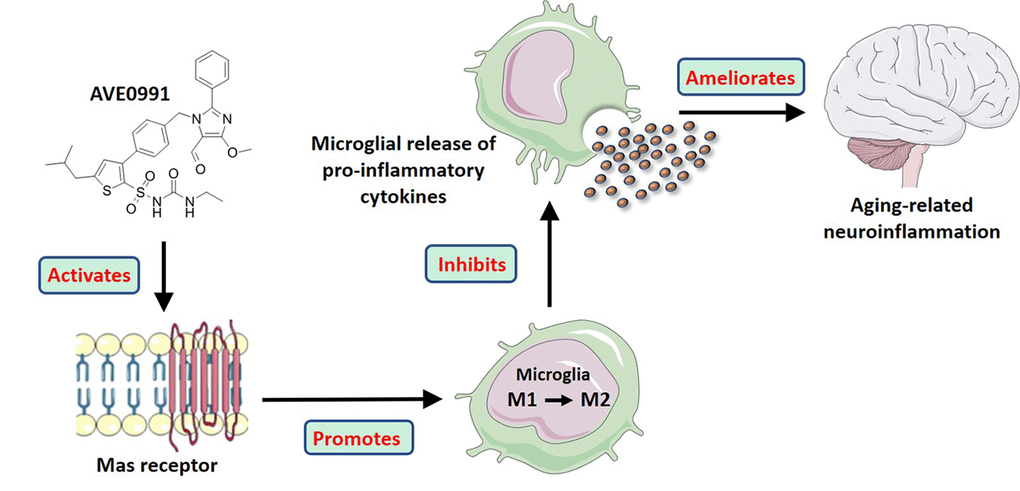
Figure 7. A model illustrating the effects of AVE0991 on aging-related neuroinflammation and its potential mechanisms.
Methods
Reagents
AVE0991, a nonpeptide agonist of MAS1 receptor [22], was purchased from Medchem Express Inc. A-779, a selective antagonist of MAS1 receptor, was purchased from Abcam Inc.
Animals and treatments
To avoid the interference of estrogen on neuroinflammation, only male animals were used in this study [32-34]. Four-, eight- and twelve-month-old male SAMP8 mice and their age-matched senescence-accelerated mouse resistant 1 (SAMR1) control mice were purchased from the Institute of Zoology, Chinese Academy of Sciences. They were maintained in individually ventilated cages in a standard animal room with a 12 h light/dark cycle and given free access to food and water as described [35]. This study was reported in accordance with the ARRIVE guideline, and the procedures in this study involving animals were in accordance with the ethical standards of Nanjing First Hospital.
Eight-month-old male SAMP8 mice were randomly allocated to five groups using a random number table generated by SPSS software, and received treatments as shown in Supplementary Figure S1. During treatment, we monitored the general health of mice and did not observe any adverse effects or significant changes in their systolic blood pressure, body weights or food intakes (data not shown).
Primary microglia cultures and treatments
Microglia were isolated from the brains of 8-month-old SAMP8 mice using a protocol described by Bliederhaeuser and colleagues with our minor modification [36-38]. Briefly, mice were deeply anesthetized using an overdose of chloral hydrate and perfused with ice-cold Ringers solution. Brains were fragmented with a scalpel and incubated with an enzymatic solution containing papain. The papain solution was quenched with 20% (FBS) in Hank’s balanced salt solution (HBSS) and centrifuged for 4 min at 200g. The pellet was resuspended in 2 mL of 0.5 mg/mL DNase in HBSS and incubated for 5 min. After being filtered through a 70-μm cell strainer and centrifuged at 200g for 4 min, the resulting pellet was then resuspended in 20% isotonic Percoll® (Sigma-Aldrich, Inc.) in HBSS. Pure HBSS was carefully laid on the top of the “percoll” layer and centrifugation was performed at 200g for 20 min. The interphase layer containing myelin and cell debris was discarded, and the pellet containing the mixed glial cell population was washed once with HBSS and suspended in Dulbecco’s modified Eagle’s medium supplemented with 10% FBS, 1%P/S, 1×pyruvate, 1×GlutaMAX™ (Thermo Fisher Scientific, Inc.) as well as 5 ng/mL of murine recombinant granulocyte and macrophage colony stimulating factor (GM-CSF). The cell suspension was plated on a 15 cm2 plate coated with poly-l-lysine and maintained in culture at 37°C, 5% CO2. The medium was replaced every 3 days until the cells reached confluency (after approximately 14-21 days in vitro (DIV)). After the glial layer becomes confluent, microglia form a nonadherent, floating cell layer. After collecting the floating layer, microglia were incubated for 3 days without GM-CSF before analysis or receiving treatments (17-24 DIV). The purity of microglia was confirmed by immunocytochemistry.
Microglia were treated for 4 h with 100 ng/ml LPS with or without 4 h pre-incubation with AVE0991 (1x10-8, 1x10-7 or 1x10-6 M) in the presence or absence of A-779 (1x10-6 M). During these treatments, cell viability was not significantly affected (data not shown). Afterwards, cells were collected for the subsequent analyses.
Double immunofluorescence assay
Double immunofluorescence assay was performed as described [39,40]. Microglia were fixed with 75% methanol in water for 10 min at -20 °C, washed with 1 mL phosphate buffer saline (PBS), and permeabilized with 0.5 mL 0.2% Triton X-100 in PBS for 15 min. Prior to the addition of antibody, cells were blocked in a blocking buffer containing PBS, 0.1% Triton X-100, and 4% bovine serum albumin (BSA) overnight. A mouse monoclonal antibody against MAS1 (1:50, Santa Cruz Biotechnology, Inc. USA) and a rabbit monoclonal antibody against AIF1 (1:100, Abcam Inc. USA) were added to the solution and incubated with the cells overnight. After washing thrice with PBS (pH 7.5) for 10 min, the cells were incubated with a TRITC-conjugated anti-mouse IgG secondary antibody (Zhongshan Goldenbridge Biotechnology, China) and a FITC-conjugated anti-rabbit IgG secondary antibody (Zhongshan Goldenbridge Biotechnology, China) in PBS with 0.1% Triton X-100 and 1% BSA for 1 h at room temperature and were viewed using a Nikon fluorescence microscope.
Brain tissue preparation
After indicated treatment, mice were killed under deep anesthesia through an overdose of chloral hydrate and were perfused transcardially with 0.9% saline (pH 7.4). Brains were then removed and stored in liquid nitrogen until use.
Quantitative Reverse Transcription PCR (qRT-PCR)
Total RNA in the whole brains or isolated mouse primary microglia was extracted by Trizol reagent as described [37,41]. Equal amounts of total RNA were reverse transcribed under standard conditions using the PrimeScript™ RT Master Mix (Takara Bio, Inc.). After that, qRT-PCR reactions were performed with SYBR Green Premix Ex Taq (Takara Bio, Inc.) and specific primers (See Supplementary Table S1).
Measurement of brain ACE2 activity
ACE2 activity in mice brain was measured by a SensoLyte 390 ACE2 activity assay kit (AnaSpec) with Mc-Ala/Dnp fluorescence resonance energy transfer peptides as described [27]. The fluorescence of Mc-Ala was monitored at excitation/emission 330/390 nm. Reactions were performed in duplicate in 96-well, clear, flat-bottom polystyrene microplates at a final volume of 100 μL. Specificity was confirmed using DX600, a specific ACE2 inhibitor (AnaSpec).
Enzyme-linked immunosorbent assay (ELISA)
The concentrations of Ang-(1-7) and inflammatory markers including IL-1β, IL-6 and TNF-α in mice brain were measured by specific ELISA kits (R&D Systems, Inc.) as described [42,43]. The alteration in absorbance in every well at 450 nm was detected with a microplate reader.
Statistical analysis
Data were analyzed using GraphPad Prism 6 (GraphPad Software, Inc.). Difference between mean values was determined by one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. Data are expressed as mean ± SD. P<0.05 was considered statistically significant.
Supplementary Materials
Conflicts of Interest
The authors have no conflicts of interest to declare.
Funding
This work was supported by National Natural Science Foundation of China (81771140), Natural Science Foundation of Jiangsu Province (BK20151084), Key Research and Development Project of Jiangsu Province (BL2014014), “Six Talent Summit” Foundation of Jiangsu Province (2016-WSN-180), Youth Medical Talent Program of Jiangsu Province (QNRC2016068, QNRC2016079), Medical Innovation Team of Jiangsu Province (CXTDA2017030), and Nanjing Medical Science and Technology Development Foundation for Distinguished Young Scholars (JQX17008).
References
- 1. Xia S, Zhang X, Zheng S, Khanabdali R, Kalionis B, Wu J, Wan W, Tai X. An Update on Inflamm-Aging: Mechanisms, Prevention, and Treatment. J Immunol Res. 2016; 2016:8426874. https://doi.org/10.1155/2016/8426874 [PubMed]
- 2. Ransohoff RM. How neuroinflammation contributes to neurodegeneration. Science. 2016; 353:777–83. https://doi.org/10.1126/science.aag2590 [PubMed]
- 3. Calsolaro V, Edison P. Neuroinflammation in Alzheimer’s disease: current evidence and future directions. Alzheimers Dement. 2016; 12:719–32. https://doi.org/10.1016/j.jalz.2016.02.010 [PubMed]
- 4. Nolan YM, Sullivan AM, Toulouse A. Parkinson’s disease in the nuclear age of neuroinflammation. Trends Mol Med. 2013; 19:187–96. https://doi.org/10.1016/j.molmed.2012.12.003 [PubMed]
- 5. Zuo Z, Qi F, Yang J, Wang X, Wu Y, Wen Y, Yuan Q, Zou J, Guo K, Yao ZB. Immunization with Bacillus Calmette-Guérin (BCG) alleviates neuroinflammation and cognitive deficits in APP/PS1 mice via the recruitment of inflammation-resolving monocytes to the brain. Neurobiol Dis. 2017; 101:27–39. https://doi.org/10.1016/j.nbd.2017.02.001 [PubMed]
- 6. García C, Gómez-Cañas M, Burgaz S, Palomares B, Gómez-Gálvez Y, Palomo-Garo C, Campo S, Ferrer-Hernández J, Pavicic C, Navarrete C, Luz Bellido M, García-Arencibia M, Ruth Pazos M, et al. Benefits of VCE-003.2, a cannabigerol quinone derivative, against inflammation-driven neuronal deterioration in experimental Parkinson’s disease: possible involvement of different binding sites at the PPARγ receptor. J Neuroinflammation. 2018; 15:19. https://doi.org/10.1186/s12974-018-1060-5 [PubMed]
- 7. Sousa C, Biber K, Michelucci A. Cellular and Molecular Characterization of Microglia: A Unique Immune Cell Population. Front Immunol. 2017; 8:198. https://doi.org/10.3389/fimmu.2017.00198 [PubMed]
- 8. Harry GJ. Microglia during development and aging. Pharmacol Ther. 2013; 139:313–26. https://doi.org/10.1016/j.pharmthera.2013.04.013 [PubMed]
- 9. Mosher KI, Wyss-Coray T. Microglial dysfunction in brain aging and Alzheimer’s disease. Biochem Pharmacol. 2014; 88:594–604. https://doi.org/10.1016/j.bcp.2014.01.008 [PubMed]
- 10. Brown GC, Vilalta A. How microglia kill neurons. Brain Res. 2015; 1628:288–97. https://doi.org/10.1016/j.brainres.2015.08.031 [PubMed]
- 11. Bessis A, Béchade C, Bernard D, Roumier A. Microglial control of neuronal death and synaptic properties. Glia. 2007; 55:233–38. https://doi.org/10.1002/glia.20459 [PubMed]
- 12. Nguyen Dinh Cat A, Touyz RM. A new look at the renin-angiotensin system--focusing on the vascular system. Peptides. 2011; 32:2141–50. https://doi.org/10.1016/j.peptides.2011.09.010 [PubMed]
- 13. Bader M, Ganten D. Update on tissue renin-angiotensin systems. J Mol Med (Berl). 2008; 86:615–21. https://doi.org/10.1007/s00109-008-0336-0 [PubMed]
- 14. Bader M. Tissue renin-angiotensin-aldosterone systems: targets for pharmacological therapy. Annu Rev Pharmacol Toxicol. 2010; 50:439–65. https://doi.org/10.1146/annurev.pharmtox.010909.105610 [PubMed]
- 15. Simões e Silva AC, Silveira KD, Ferreira AJ, Teixeira MM. ACE2, angiotensin-(1-7) and Mas receptor axis in inflammation and fibrosis. Br J Pharmacol. 2013; 169:477–92. https://doi.org/10.1111/bph.12159 [PubMed]
- 16. Khajah MA, Fateel MM, Ananthalakshmi KV, Luqmani YA. Anti-Inflammatory Action of Angiotensin 1-7 in Experimental Colitis. PLoS One. 2016; 11:e0150861. https://doi.org/10.1371/journal.pone.0150861 [PubMed]
- 17. Meng Y, Li T, Zhou GS, Chen Y, Yu CH, Pang MX, Li W, Li Y, Zhang WY, Li X. The angiotensin-converting enzyme 2/angiotensin (1-7)/Mas axis protects against lung fibroblast migration and lung fibrosis by inhibiting the NOX4-derived ROS-mediated RhoA/Rho kinase pathway. Antioxid Redox Signal. 2015; 22:241–58. https://doi.org/10.1089/ars.2013.5818 [PubMed]
- 18. Feltenberger JD, Andrade JM, Paraíso A, Barros LO, Filho AB, Sinisterra RD, Sousa FB, Guimarães AL, de Paula AM, Campagnole-Santos MJ, Qureshi M, dos Santos RA, Santos SH. Oral formulation of angiotensin-(1-7) improves lipid metabolism and prevents high-fat diet-induced hepatic steatosis and inflammation in mice. Hypertension. 2013; 62:324–30. https://doi.org/10.1161/HYPERTENSIONAHA.111.00919 [PubMed]
- 19. Jiang T, Gao L, Guo J, Lu J, Wang Y, Zhang Y. Suppressing inflammation by inhibiting the NF-κB pathway contributes to the neuroprotective effect of angiotensin-(1-7) in rats with permanent cerebral ischaemia. Br J Pharmacol. 2012; 167:1520–32. https://doi.org/10.1111/j.1476-5381.2012.02105.x [PubMed]
- 20. Tomobe K, Nomura Y. Neurochemistry, neuropathology, and heredity in SAMP8: a mouse model of senescence. Neurochem Res. 2009; 34:660–69. https://doi.org/10.1007/s11064-009-9923-x [PubMed]
- 21. Akiguchi I, Pallàs M, Budka H, Akiyama H, Ueno M, Han J, Yagi H, Nishikawa T, Chiba Y, Sugiyama H, Takahashi R, Unno K, Higuchi K, Hosokawa M. SAMP8 mice as a neuropathological model of accelerated brain aging and dementia: toshio Takeda’s legacy and future directions. Neuropathology. 2017; 37:293–305. https://doi.org/10.1111/neup.12373 [PubMed]
- 22. Wiemer G, Dobrucki LW, Louka FR, Malinski T, Heitsch H. AVE 0991, a nonpeptide mimic of the effects of angiotensin-(1-7) on the endothelium. Hypertension. 2002; 40:847–52. https://doi.org/10.1161/01.HYP.0000037979.53963.8F [PubMed]
- 23. Liu M, Shi P, Sumners C. Direct anti-inflammatory effects of angiotensin-(1-7) on microglia. J Neurochem. 2016; 136:163–71. https://doi.org/10.1111/jnc.13386 [PubMed]
- 24. Franco R, Fernández-Suárez D. Alternatively activated microglia and macrophages in the central nervous system. Prog Neurobiol. 2015; 131:65–86. https://doi.org/10.1016/j.pneurobio.2015.05.003 [PubMed]
- 25. Griñan-Ferré C, Palomera-Ávalos V, Puigoriol-Illamola D, Camins A, Porquet D, Plá V, Aguado F, Pallàs M. Behaviour and cognitive changes correlated with hippocampal neuroinflammaging and neuronal markers in female SAMP8, a model of accelerated senescence. Exp Gerontol. 2016; 80:57–69. https://doi.org/10.1016/j.exger.2016.03.014 [PubMed]
- 26. Jiang T, Yu JT, Zhu XC, Tan MS, Gu LZ, Zhang YD, Tan L. Triggering receptor expressed on myeloid cells 2 knockdown exacerbates aging-related neuroinflammation and cognitive deficiency in senescence-accelerated mouse prone 8 mice. Neurobiol Aging. 2014; 35:1243–51. https://doi.org/10.1016/j.neurobiolaging.2013.11.026 [PubMed]
- 27. Jiang T, Zhang YD, Zhou JS, Zhu XC, Tian YY, Zhao HD, Lu H, Gao Q, Tan L, Yu JT. Angiotensin-(1-7) is Reduced and Inversely Correlates with Tau Hyperphosphorylation in Animal Models of Alzheimer’s Disease. Mol Neurobiol. 2016; 53:2489–97. https://doi.org/10.1007/s12035-015-9260-9 [PubMed]
- 28. Jiang T, Gao L, Lu J, Zhang YD. ACE2-Ang-(1-7)-Mas Axis in Brain: A Potential Target for Prevention and Treatment of Ischemic Stroke. Curr Neuropharmacol. 2013; 11:209–17. https://doi.org/10.2174/1570159X11311020007 [PubMed]
- 29. Marshall AC, Shaltout HA, Pirro NT, Rose JC, Diz DI, Chappell MC. Enhanced activity of an angiotensin-(1-7) neuropeptidase in glucocorticoid-induced fetal programming. Peptides. 2014; 52:74–81. https://doi.org/10.1016/j.peptides.2013.12.006 [PubMed]
- 30. Domenig O, Manzel A, Grobe N, Kaltenecker C, Kovarik J, Stegbauer J, Gurley SB, Antlanger M, Elased K, Saemann M, Linker R, Poglitsch M. 8d.05: The Role Of Neprilysin In Angiotensin 1-7 Formation In the Kidney. J Hypertens. 2015 (Suppl 1); 33:e114–15. https://doi.org/10.1097/01.hjh.0000467659.31038.0f [PubMed]
- 31. Olszanecki R, Madej J, Suski M, Gebska A, Bujak-Gizycka B, Korbut R. Angiotensin metabolism in rat stomach wall: prevalence of angiotensin-(1-7) formation. J Physiol Pharmacol. 2009; 60:191–96. [PubMed]
- 32. Wei J, Yuen EY, Liu W, Li X, Zhong P, Karatsoreos IN, McEwen BS, Yan Z. Estrogen protects against the detrimental effects of repeated stress on glutamatergic transmission and cognition. Mol Psychiatry. 2014; 19:588–98. https://doi.org/10.1038/mp.2013.83 [PubMed]
- 33. Benedusi V, Meda C, Della Torre S, Monteleone G, Vegeto E, Maggi A. A lack of ovarian function increases neuroinflammation in aged mice. Endocrinology. 2012; 153:2777–88. https://doi.org/10.1210/en.2011-1925 [PubMed]
- 34. Bjorling DE, Wang ZY. Estrogen and neuroinflammation. Urology. 2001 (Suppl 1); 57:40–46. https://doi.org/10.1016/S0090-4295(01)01124-4 [PubMed]
- 35. Tan MS, Yu JT, Jiang T, Zhu XC, Guan HS, Tan L. IL12/23 p40 inhibition ameliorates Alzheimer’s disease-associated neuropathology and spatial memory in SAMP8 mice. J Alzheimers Dis. 2014; 38:633–46. https://doi.org/10.3233/JAD-131148 [PubMed]
- 36. Bliederhaeuser C, Grozdanov V, Speidel A, Zondler L, Ruf WP, Bayer H, Kiechle M, Feiler MS, Freischmidt A, Brenner D, Witting A, Hengerer B, Fändrich M, et al. Age-dependent defects of alpha-synuclein oligomer uptake in microglia and monocytes. Acta Neuropathol. 2016; 131:379–91. https://doi.org/10.1007/s00401-015-1504-2 [PubMed]
- 37. Jiang T, Zhang YD, Gao Q, Zhou JS, Zhu XC, Lu H, Shi JQ, Tan L, Chen Q, Yu JT. TREM1 facilitates microglial phagocytosis of amyloid beta. Acta Neuropathol. 2016; 132:667–83. https://doi.org/10.1007/s00401-016-1622-5 [PubMed]
- 38. Jiang T, Wan Y, Zhang YD, Zhou JS, Gao Q, Zhu XC, Shi JQ, Lu H, Tan L, Yu JT. TREM2 Overexpression has No Improvement on Neuropathology and Cognitive Impairment in Aging APPswe/PS1dE9 Mice. Mol Neurobiol. 2017; 54:855–65. https://doi.org/10.1007/s12035-016-9704-x [PubMed]
- 39. Chen SS, Jiang T, Wang Y, Gu LZ, Wu HW, Tan L, Guo J. Activation of double-stranded RNA-dependent protein kinase inhibits proliferation of pancreatic β-cells. Biochem Biophys Res Commun. 2014; 443:814–20. https://doi.org/10.1016/j.bbrc.2013.12.051 [PubMed]
- 40. Jiang T, Gao L, Zhu XC, Yu JT, Shi JQ, Tan MS, Lu J, Tan L, Zhang YD. Angiotensin-(1-7) inhibits autophagy in the brain of spontaneously hypertensive rats. Pharmacol Res. 2013; 71:61–68. https://doi.org/10.1016/j.phrs.2013.03.001 [PubMed]
- 41. Jiang T, Zhang YD, Chen Q, Gao Q, Zhu XC, Zhou JS, Shi JQ, Lu H, Tan L, Yu JT. TREM2 modifies microglial phenotype and provides neuroprotection in P301S tau transgenic mice. Neuropharmacology. 2016; 105:196–206. https://doi.org/10.1016/j.neuropharm.2016.01.028 [PubMed]
- 42. Jiang T, Yu JT, Zhu XC, Zhang QQ, Tan MS, Cao L, Wang HF, Lu J, Gao Q, Zhang YD, Tan L. Angiotensin-(1-7) induces cerebral ischaemic tolerance by promoting brain angiogenesis in a Mas/eNOS-dependent pathway. Br J Pharmacol. 2014; 171:4222–32. https://doi.org/10.1111/bph.12770 [PubMed]
- 43. Jiang T, Tan L, Zhu XC, Zhou JS, Cao L, Tan MS, Wang HF, Chen Q, Zhang YD, Yu JT. Silencing of TREM2 exacerbates tau pathology, neurodegenerative changes, and spatial learning deficits in P301S tau transgenic mice. Neurobiol Aging. 2015; 36:3176–86. https://doi.org/10.1016/j.neurobiolaging.2015.08.019 [PubMed]