



Introduction
Gastric cancer (GC) accounts for over 10% of new cancer cases each year [1–3]. It is one leading cause of cancer-associated human mortalities [2, 3]. The prognosis of advanced, recurrent and metastatic GC is far from satisfactory. Current treatment options for this devastating disease are extremely limited [2, 3]. Molecularly-targeted therapies are the research focus for better GC therapies [4, 5]. Identification of novel therapeutic targets and biomarkers of GC is extremely urgent [4, 5].
LIM homeobox transcription factor 1, alpha (LMX1A) is a widely-studied member of LIM-homeodomain (LIM-HD) family protein [6]. As an evolutionary conserved transcription factor, LMX1A regulates a number of key physiological and pathological processes [6]. LMX1A functions as a tumor suppressor, downregulated in GC and many other cancers [7–13]. LMX1A gene promoter hyper-methylation in human cancers could be one important cause of its downregulation [7, 8, 10]. Our previous study has identified the LMX1A-targeting miRNA, microRNA-9 (miR-9) [9]. miR-9 upregulation might account for LMX1A downregulation in human GC tissues [9]. Furthermore, miR-9 inhibition upregulated its target LMX1A, thereby inhibiting GC cell survival and proliferation [9]. The underlying mechanism of miR-9 upregulation in human GC is still largely unknown.
Long non-coding RNAs (LncRNAs) are a family of evolutionarily conserved non-coding RNAs (ncRNAs) with over 200 nucleotide long [14–16]. Dysregulation of LncRNAs is commonly detected in GC [17, 18], which is associated with cancer progression and patients’ prognosis [14, 17–20]. LncRNAs regulate almost all important cellular functions, from genomic imprinting, cell proliferation and growth, cell cycle progression to cell differentiation, survival and apoptosis [14–16]. LncRNA acts as competing endogenous RNA (ceRNA) to sponge target miRNAs [17, 21]. The results of the present study will show that LINC00682 (long intergenic non-protein coding RNA 682) targets miR-9-LMX1A signaling axis to inhibit human GC cell survival and proliferation.
Results
Ectopic overexpression of LINC00682 induces miR-9 downregulation but LMX1A upregulation, inhibiting AGS cell survival, proliferation, migration and invasion
We hypothesized that miR-9 upregulation in GC tissues (see our previous study [9]) could possibly be due to downregulation of certain LncRNAs. Therefore, LncBase (Predicted v.2) was searched to find possible miR-9-targeting LncRNAs. The LncRNAs were further verified by searching other LncRNA/miRNA databases (StarBase and miRbase). The bioinformatic analyses identified that one LncRNA, LINC00682, putatively targets miR-9, with its percentage over 99%.
In order to study the potential effect of LINC00682 on miR-9-LMX1A axis, the lentivirus encoding LINC00682-expressing construct (“LV-LINC00682”) was added to AGS cells. Following selection by the puromycin-containing complete medium, two stable cell lines, “sLi-1” and “sLi-2”, were established. qPCR results confirmed that LINC00682 levels increased over ten folds (versus control cells) in the LV-LINC00682-expressing stable cells (Figure 1A). Importantly, LINC00682 overexpression in AGS cells induced significant downregulation of miR-9 (Figure 1B), but a significant increase in LMX1AUTR luciferase activity (Figure 1C). Consequently, LMX1A mRNA levels increased over five-six folds by LV-LINC00682 (Figure 1D). Western blotting results confirmed that forced overexpression of LINC00682 induced LMX1A protein upregulation as well (Figure 1E). LMX1B mRNA and protein expression was however not significantly affected by LV-LINC00682 (Figure 1E and 1F).
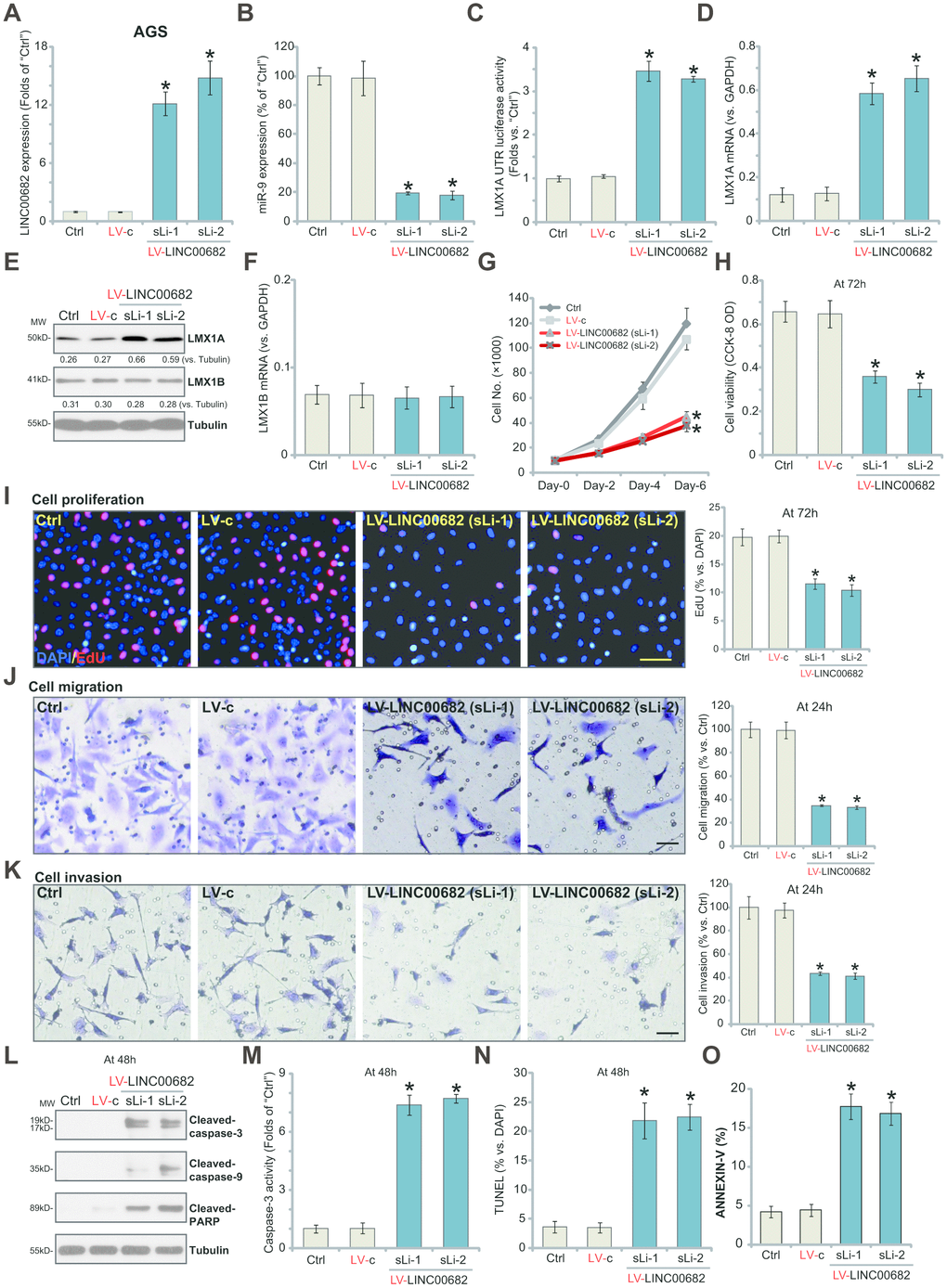
Figure 1. Ectopic overexpression of LINC00682 induces miR-9 downregulation but LMX1A upregulation, inhibiting AGS cell survival, proliferation, migration and invasion. AGS cells were infected with LINC00682-expressing lentivirus (“LV-LINC00682”), following puromycin selection two stable lines (“sLi-1/-2”) were obtained; Control cells were infected with the lentivirus with empty vector (“LV-c”); In those cells expression of LINC00682 (A), miR-9 (B), LMX1A mRNA (D), LMX1B mRNA (F) was tested by qPCR; The relative LMX1A3’-UTR luciferase activity was tested (C); Expression of the listed proteins in total cell lysates was tested by Western blotting (E); Cells were further cultured for the indicated time periods, cell survival, proliferation, migration and invasion in vitro were tested by the appropriate assays (G–K); Cell apoptosis was tested by Western blotting assay of apoptosis proteins (L), caspase-3 activity assay (M), nuclear TUNEL staining assay (N) and Annexin V FACS staining (O). The exact same number of viable cells of different genetic treatments were plated initially (“0h”/“Day-0”) for the functional assays (Same for all following Figures). Five repeated views in each condition were included to calculate the average number of migrated/invasive cells (Same for all Figures). Listed proteins were quantified and normalized to the loading control (E). “MW” stands for molecular weight (Same for all Figures). “Ctrl” stands for the parental control cells (Same for all Figures). For each assay, n=5 (five dishes or wells). *P <0.05 vs. “LV-c” cells. Experiments in this figure were repeated four times, and similar results were obtained. Bar=100 μm (I, J and K).
Our previous study has demonstrated that LMX1A functions as a tumor suppressor, inhibiting GC cell survival and proliferation [9]. By counting cell number, we show that forced overexpression of LINC00682 by LV-LINC00682 significantly inhibited AGS cell growth (Figure 1G). Furthermore, AGS cells with LV-LINC00682 presented with decreased cell viability (CCK-8 OD, Figure 1H) and inhibited EdU ratio (Figure 1I), suggesting proliferation inhibition. Testing cell migration, by the “Transwell” assays, show that LV-LINC00682-induced LINC00682 overexpression significantly inhibited AGS cell migration in vitro (Figure 1J). Furthermore, the “Matrigel Transwell” assay results demonstrated that AGS cell invasion was also suppressed by ectopic LINC00682 overexpression (Figure 1K).
Importantly, significant apoptosis activation was detected in LINC00682-overexpressed AGS cells, evidenced by cleavages of caspase-3, caspase-9 and poly (ADP-ribose) polymerase (PARP) (Figure 1L), as well as increased caspase-3 activation (Figure 1M) and nuclear TUNEL ratio (Figure 1N). Additionally, LINC00682-overexpressed AGS cells presented with increased Annexin V staining (Figure 1O). The control lentivirus with empty vector (“LV-c”) had no significant effect on LINC00682-miR-9-LMX1A/B expression (Figure 1A–1F) nor AGS cell functions (Figure 1G–1O). These results show that ectopic overexpression of LINC00682 induces miR-9 downregulation but LMX1A upregulation, inhibiting AGS cell survival, proliferation, migration and invasion.
LINC00682 knockdown induces miR-9 upregulation but LMX1A downregulation, promoting AGS cell survival, proliferation, migration and invasion
Since exogenous LINC00682 overexpression inhibited AGS cell progression in vitro (Figure 1), we hypothesized that LINC00682 silencing might promote cell progression. To test this hypothesis, two different siRNAs, targeting non-overlapping sequences (“Seq1/Seq2”) of LINC00682 were transfected individually to AGS cells. Results from the qPCR confirmed that each siRNA resulted in over 90% reduction of LINC00682 expression in AGS cells (Figure 2A). miR-9 levels were significantly increased in LINC00682-silenced cells (Figure 2B), where LMX1A 3’-UTR luciferase activity was largely decreased (Figure 2C). In AGS cells LMX1A mRNA (Figure 2D) and protein (Figure 2E) levels were significantly downregulated by LINC00682 siRNAs. While the two had no effect on LMX1B expression (Figure 2E and 2F).
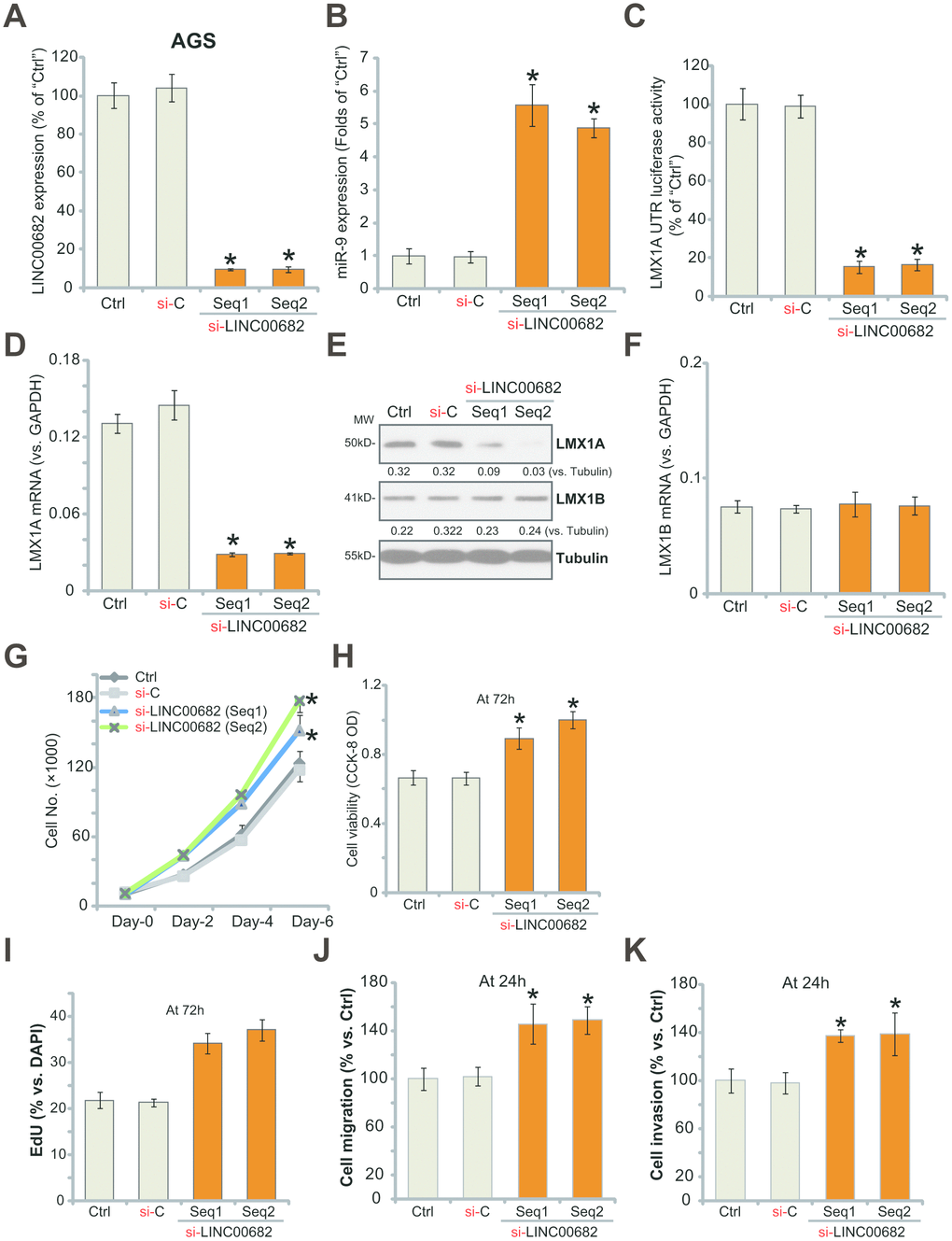
Figure 2. LINC00682 knockdown induces miR-9 upregulation but LMX1A downregulation, promoting AGS cell survival, proliferation, migration and invasion. AGS cells were transfected with 500 nM of LINC00682 siRNA (“Seq1/Seq2”, two rounds, total 48h) or the scramble non-sense control siRNA (“si-C”), expression levels of LINC00682 (A), miR-9 (B), LMX1A mRNA (D), LMX1B mRNA (F) were tested by qPCR; The relative LMX1A3’-UTR luciferase activity was tested (C); Expression of the listed proteins in total cell lysates was tested by Western blotting (E); Cells were further cultured for the indicated time periods, cell survival and proliferation in vitro were tested by the appropriate assays (G-I); Cell migration and invasion were tested by “Transwell” (J) and “Matrigel Transwell” (K) assays, respectively. Listed proteins were quantified and normalized to the loading control (E). For each assay, n=5 (five dishes or wells). *P <0.05 vs. “si-C” cells. Experiments in this figure were repeated four times, and similar results were obtained.
For the functional studies, LINC00682 silencing increased AGS cell growth (Figure 2G), cell viability (CCK-8 OD, Figure 2H), nuclear EdU staining (Figure 2I). Results from “Transwell” and “Matrigel Transwell” assays demonstrated that LINC00682 silencing by targeted siRNAs promoted AGS cell migration (Figure 2J) and invasion (Figure 2K) in vitro. The non-sense scramble control siRNA (“si-C”) had no detectable effect on LINC00682-miR-9-LMX1A/B expression (Figure 2A–2F) nor AGS cell functions (Figure 2G–2K). Therefore, LINC00682 knockdown promoted AGS cell progression in vitro.
Ectopic overexpression of LINC00682 induces miR-9 downregulation but LMX1A upregulation, inhibiting survival and proliferation of primary human GC cells
The potential effect of LINC00682 on the primary GC cells was studied next. As reported [9], the primary human GC cells were derived from three different primary GC patients (“GC-1/GC-2/GC-3”) and cultured in vitro. The primary cells were infected with LV-LINC00682 (see Figure 1), followed by selection in puromycin-containing medium. Control cells were treated with the control lentivirus (“LV-c”, see Figure 1). qPCR results in Figure 3A confirmed that LINC00682 levels were significantly increased (over eight to ten folds) in stable primary cancer cells with LV-LINC00682. Forced overexpression of LINC00682 induced miR-9 downregulation (Figure 3B), while upregulating LMX1A mRNA (Figure 3C) and protein (Figure 3D) in the primary GC cells.
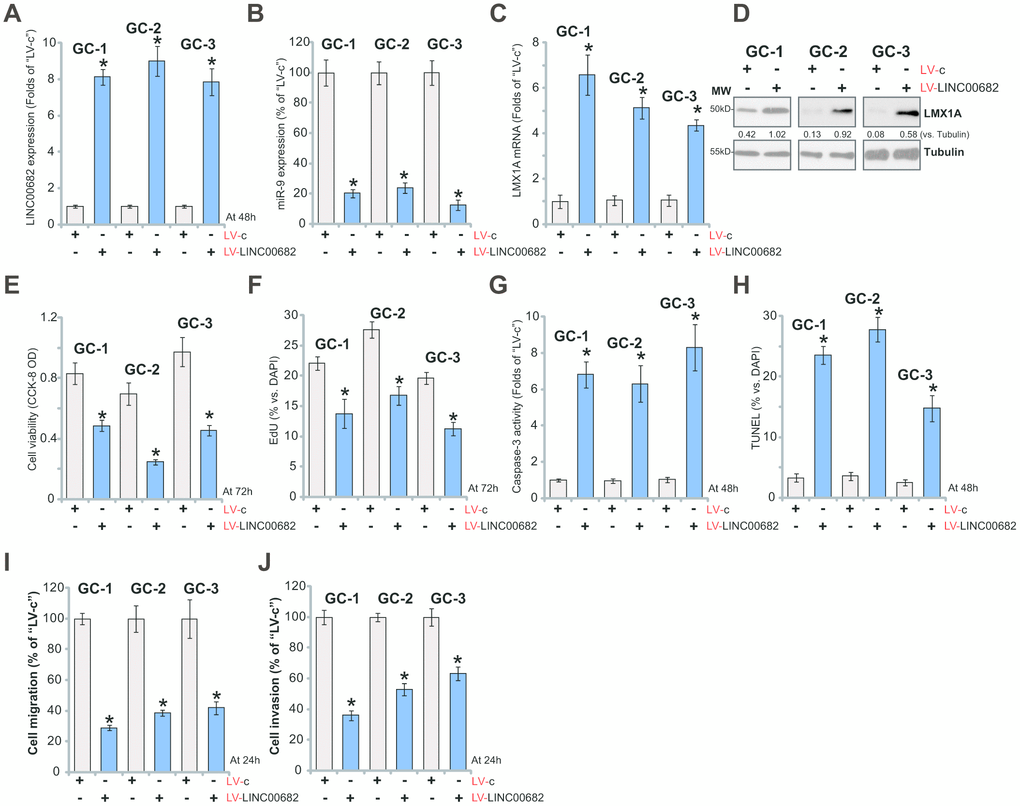
Figure 3. Ectopic overexpression of LINC00682 induces miR-9 downregulation but LMX1A upregulation, inhibiting survival and proliferation of primary human GC cells. The primary human GC cells, derived from three different primary GC patients (“GC-1/GC-2/GC-3”), were infected with LINC00682-expressing lentivirus (“LV-LINC00682”) or the lentivirus with empty vector (“LV-c”), followed by puromycin selection; Expression levels of LINC00682 (A), miR-9 (B), LMX1A mRNA (C), and listed proteins (D) were tested; Cells were further cultured for the indicated time periods, cell viability (CCK-8 OD, E) and proliferation (EdU staining, F) were tested; Cell apoptosis was tested by caspase-3 activity assay (G) and TUNEL staining (H). Cell migration and invasion were tested by “Transwell” (I) and “Matrigel Transwell” (J) assays, respectively. Listed proteins were quantified and normalized to the loading control (D). For each assay, n=5 (five dishes or wells). *P <0.05 vs. “LV-c” cells. Experiments in this figure were repeated three times, and similar results were obtained.
When studying cellular functions, we show that LV-LINC00682 inhibited cell viability (Figure 3E) and EdU staining (Figure 3F) in primary GC cells. On the other hand, cell apoptosis, tested by caspase-3 activation (Figure 3G) and TUNEL ratio increase (Figure 3H), was induced by LV-LINC00682. “Transwell” and “Matrigel Transwell” assay results, Figure 3I and 3J, demonstrated that ectopic LINC00682 overexpression inhibited in vitro migration and invasion of the primary human GC cells. These results show that LINC00682 overexpression inhibited primary GC cell survival and proliferation, migration and invasion, while provoking apoptosis activation.
LINC00682 inhibits AGS cell progression via targeting miR-9-LMX1A axis
If miR-9 is the primary target of LINC00682, restoring miR-9 expression should abolish LINC00682-induced actions in GC cells. Thus, in the stable AGS cells with LV-LINC00682 (“sLi-1”, see Figure 1), the miR-9-expressing lentivirus (“lv-miR-9”, see our previous study [9]) was added. Two stable cell lines were established, “sL1/sL2”. As shown, in LV-LINC00682 AGS cells, lv-miR-9 did not affect LINC00682 expression (Figure 4A). Yet it restored miR-9 expression, four-five times higher to the control level (Figure 4B). Further, LV-LINC00682-induced LMX1A mRNA (Figure 4C) and protein (Figure 4D) upregulation was completely blocked by lv-miR-9. LMX1B mRNA was again unchanged (Figure 4E). Importantly, LV-LINC00682-induced viability (CCK-8 OD) reduction (Figure 4F) and apoptosis activation (the increase in TUNEL staining) (Figure 4G) were abolished by lv-miR-9 in AGS cells. These results showed that ectopic miR-9 expression reversed LINC00682-induced inhibition on GC cells, suggesting that miR-9 is the target of LINC00682.
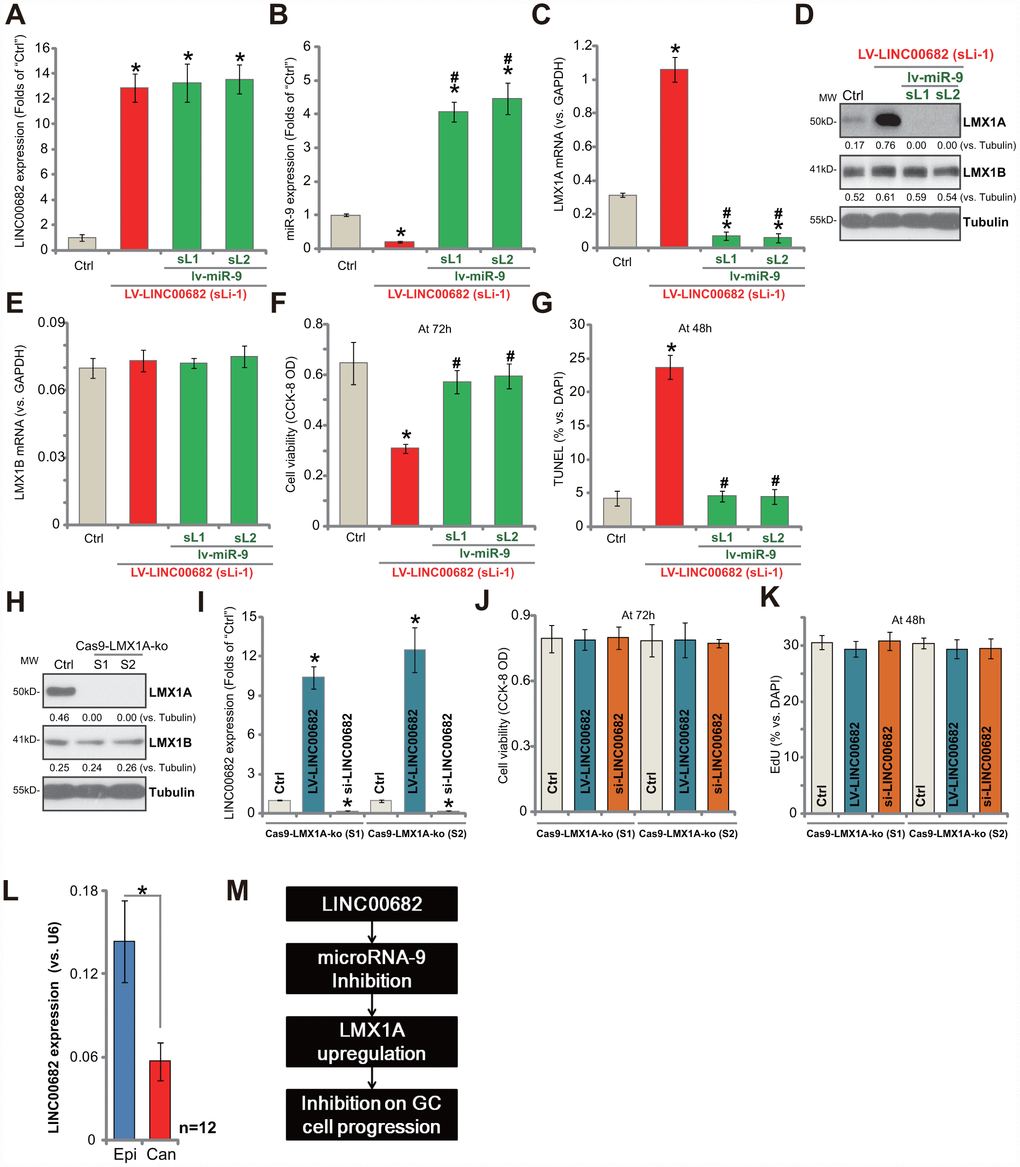
Figure 4. LINC00682 inhibits AGS cell progression via targeting miR-9-LMX1A axis. AGS cells were infected with LINC00682-expressing lentivirus (“LV-LINC00682”), following puromycin selection the stable cells were established. The stable cells (“sLi-1”) were further infected with pri-miR-9-expressing lentivirus (“lv-miR-9”) for 24h, following puromycin selection two stable lines were obtained (“sL1/ sL2”); In the cells expression of LINC00682 (A), miR-9 (B), LMX1A mRNA (C), listed proteins (D) and LMX1B mRNA (E) was tested; Cells were further cultured for applied time, and cell viability (F) and apoptosis (G) were tested by the appropriate assays. AGS cells were transfected with the lenti-CRISPR/Cas9 LMX1A knockout constructs with non-overlapping sgRNA sequences (“S1/S2”), following FACS sorting and puromycin selection two stable lines were obtained (“Cas9-LMX1A-ko”). LMX1A and LMX1B expression was tested (H). LV-LINC00682 or LINC00682 siRNA (500 nM) were transfected to the Cas9-LMX1A-ko AGS cells (“S1/S2”) for 72h, LINC00682 expression (I), cell viability (J) and proliferation (K) were tested. Expression of LINC00682 in twelve (n=12) human GC tissues (“Can”) and matched surrounding normal epithelial tissues (“Epi”) was tested by qPCR, and results were normalized to U6 RNA (L). The proposed signaling pathway of this study (M). Listed proteins were quantified and normalized to the loading control (D and H). For each assay, n=5 (five dishes or wells, except for L). *P <0.05 vs. “Ctrl” cells. #P <0.05 vs. cells without “lv-miR-9” (B, C, E–G). *P <0.05 (L). Experiments in this figure were repeated three times, and similar results were obtained.
Our study has previously shown that LMX1A is the direct and primary target of miR-9 in GC cells, therefore LINC00682 should be ineffective in LMX1A-depleted cells. To test this hypothesis, the CRISPR/Cas9 method by using non-overlapping sgRNA sequences (“S1/S2”) [9] was utilized to knockout LMX1A in AGS cells. Two stable LMX1A knockout (“Cas9-LMX1A-ko”) cell lines were established. Testing LMX1A protein expression by Western blotting confirmed complete LMX1A KO in the stable cells (Figure 4H). LV-LINC00682 (see Figure 1) or LINC00682 siRNA (“Seq1”, see Figure 2) were transfected to LMX1A KO AGS cells, which significantly altered LINC00682 expression (Figure 4I). Yet, neither LV-LINC00682 nor LINC00682 siRNA affected viability (CCK-8 OD, Figure 4J) and proliferation (EdU ratio, Figure 4K) in the LMX1A KO cells. Therefore, LINC00682 was completely ineffective in LMX1A KO AGS cells, confirming LMX1A is the target protein of LINC00682.
LINC00682 is downregulated in human GC tissues
Expression of LINC00682 in human GC tissues was tested. Total RNA was extracted from fresh GC tissues and paired adjacent normal epithelial tissues from twelve (12) primary GC patients [9]. LINC00682 expression was examined by qPCR. Results show that LINC00682 levels are significantly downregulated in cancer tissues (“Can”) (Figure 4L), when compared to those in the adjacent epithelial (“Epi”) tissues (Figure 4L). Therefore, LINC00682 downregulation correlates with miR-9 upregulation and LMX1A downregulation in GC tissues (see the results from same set of tissue samples [9]).
Discussion
LMX1A is hyper-methylated and downregulated in ovarian cancer and cervical cancer [12, 13]. Recent studies have proposed LMX1A as a tumor suppressor in GC and other cancers. Chao et al., have shown that LMX1A can inhibit tumorigenesis, epithelial-mesenchymal transition (EMT) and stem-like properties of epithelial ovarian cancer [11]. Its expression is associated with advanced stages, poor differentiation, early recurrence and poor overall survival in ovarian cancer [11]. Feng et al., demonstrated that LMX1A inhibited GC cell metastasis through negative regulation of β-catenin [10]. Our previous study has demonstrated that LMX1A is downregulated in human GC. Furthermore CRISPR/Cas9-mediated LMX1A KO promoted GC cell proliferation [9]. Conversely, LMX1A upregulation by miR-9 depletion led to GC cell apoptosis [9]. However, the regulation of LMX1A in carcinogenesis remains largely unknown.
Very few studies have explored the potential biological function of LINC00682. One genome wide methylation study between primary and recurrent hepatocellular carcinomas (HCC) has indicated that low levels of LINC00682 methylation were strongly correlated with HCC recurrence and patient disease/recurrent-free survival (DFS) [22]. In the present study, our results indicate that LINC00682 could function as the ceRNA of miR-9 to promote LMX1A expression, thereby inhibiting GC cell progression.
We show that ectopic overexpression of LINC00682 induced miR-9 downregulation but LMX1A upregulation, inhibiting GC cell survival, proliferation migration and invasion. Furthermore, significant apoptosis activation was detected in LINC00682-overexpressed GC cells. On the contrary, LINC00682 knockdown by targeted siRNAs induced miR-9 upregulation but LMX1A downregulation, promoting AGS cell proliferation, migration and invasion. Significantly, LINC00682 expression levels are downregulated in human GC tissues, correlated with miR-9 upregulation and LMX1A downregulation. Thus, LINC00682 downregulation could be at least one reason of miR-9-LMX1A dysregulation in human GC (see proposed signaling carton in Figure 4M).
Our results suggest that miR-9-LMX1A signaling axis is the direct target of LINC00682 in GC cells. In AGS cells, restoring miR-9 expression by lv-miR-9 reversed LV-LINC00682-induced LMX1A upregulation and cancer cell inhibition. Furthermore, we have previously shown that miR-9 inhibition by antagomir-9 increased LMX1A expression to inhibit GC cell proliferation [9]. Thus, antagomir-9 exerted similar functions as LV-LINC00682. Our results demonstrated that neither LINC00682 overexpression nor LINC00682 silencing altered the functions of LMX1A KO AGS cells. We conclude that LMX1A is the target protein of LINC00682 in GC cells.
In conclusion, LINC00682 inhibits GC cell progression via targeting miR-9-LMX1A signaling axis.
Materials and Methods
Chemicals and reagents
Puromycin and polybrene were provided by Sigma-Aldrich (St. Louis, Mo). The antibodies of the present study were obtained from Abcam (Cambridge, MA). The reagents for RNA assays, Lipofectamine 2000 and other transfection reagents were provided by Thermo-Fisher (Shanghai, China). All sequences, constructs, viruses and plasmids were provided by Shanghai Genechem Co. (Shanghai, China).
Cell culture
Using a previously described protocol AGS cells were cultured [9]. The primary human GC cells, derived from three written-informed consent GC patients (“GC-1/-2/-3”), were cultured in the described medium [23]. The enrolled primary GC patients in this study received no prior chemotherapy/radiotherapy before surgeries. The protocols were approved by the Ethics Board of Fudan University, in according to Declaration of Helsinki.
Human tissues
As described early [9], from twelve (12) informed-consent primary GC patients, the fresh human GC tissues and paired surrounding gastric epithelial tissues were acquired. Tissues were washed, minced, and homogenized in tissue lysis buffer (Biyuntian, Wuxi, China), stored in liquid nitrogen. Expression of LINC00682 was tested by quantitative reverse transcriptase PCR (“qPCR”).
qPCR assay
The detailed protocol for qPCR was described early [9]. The ΔΔCt method was utilized for the quantification of target mRNA, with GAPDH as the internal control. LINC00682 and miR-9 expression was normalized to U6 RNA. Primers for miR-9, LMX1A, GAPDH and U6 were described previously [9]. The primers for LINC00682 were provided by Shanghai Genechem (Shanghai, China).
Forced LINC00682 overexpression
The full-length LINC00682 was synthesized by Shanghai Genechem, sub-cloned to a GV248 (hU6-MCS-Ubiquitin-EGFP-IRES-puromycin) vector (Shanghai Genechem). The construct was transfected to HEK-293 cells together with lentiviral packaging plasmids [9] to generate LINC00682-expressing lentivirus (“LV-LINC00682”). After filtration and enrichment, the lentivirus was added to cultured GC cells (in polybrene medium). Thereafter, puromycin (5.0 μg/mL) was added to select stable cells for 4-5 passages. Control cells were infected with lentivirus with empty vector (“LV-c”).
LINC00682 small interfering RNA (siRNA)
Two siRNAs (provided and verified again by Shanghai Genechem) targeting non-overlapping sequences (“Seq1/2”) of LINC00682 were individually transfected by Lipofectamine 2000 for 24h (the siRNA concentration at 500 nM). The transfection was repeated another round (total 48h). Knockdown efficiency was verified by qPCR testing LINC00682 expression. Control cells were transfected with the scramble non-sense control siRNA (“si-C”).
LMX1A 3’-UTR luciferase reporter assay
As reported early [9], AGS cells were transfected with pGL4.13 LMX1A 3’-UTR construct [9], the Renillaluciferase reporter vector and pRL-SV40 (Promega). In AGS cells following transfection of LINC00682 siRNA or LV-LINC00682, LMX1A3’-UTR luciferase activity was tested as reported [9].
Cell viability
GC cells were seeded into the 96-well plates at 5000 cells per well [9]. Following the applied genetic treatments, the viability was determined by Cell Counting kit-8 (CCK-8, Dojindo Laboratories, Kumamoto, Japan). CCK-8 optical density (OD) values were tested at the wavelength of 570 nm.
In vitro cell migration and invasion assays
As described [24], GC cells were seeded on “Transwell” upper chamber (at 3 × 4000 cells per chamber, BD Biosciences). The complete medium (with 10% FBS) was added to the lower compartments. After 24h the migrated cells on the lower surface were stained. Matrigel (Sigma) was added in the chamber surface when analyzing cell invasion. Five repeated views in each condition were included to calculate the average number of migrated/invasive cells.
EdU assay
Following the genetic treatments, GC cells were seeded onto 12-well plates at 30, 000 cells per well. The EdU (5-ethynyl-20-deoxyuridine) Apollo-567 Kit (RIBOBIO, Shanghai, China) was employed to quantify cell proliferation. EdU and DAPI dyes were added to GC cells for 6h. Under a fluorescent microscope cell nuclei were visualized. For each condition 800 nuclei in five random views were included to calculate EdU ratio (EdU/DAPI×100%).
Apoptosis assays
Testing cell apoptosis, by the caspase-3 activity assay, terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL), and Propidium iodide (PI)-Annexin V FACS assay was described in other studies [9, 25].
LMX1A knockout
AGS cells were seeded into six-well plates at 100, 000 cells per well. The two lentiCRISPR-GFP-puro LMX1A knockout constructs (with non-overlapping small guide RNAs [“S1/S2”], see our previous study [9]) were individually transfected to AGS cells by Lipofectamine 2000. FACS-sorting of GFP-positive cells was performed, and resulting cells were further cultured in puromycin-containing medium. LMX1A knockout in the stable cells was verified by Western blotting.
Western blotting
Following the genetic treatments, the lysis buffer (Biyuntian, Wuxi, China) was added to cultured GC cells. Quantified total cellular lysates were separated by a SDS-PAGE (10%) gel, transferred to a polyvinylidene fluoride blot. The latter was blocked in PBST with 10% milk, and incubated with indicated primary and secondary antibodies. The immunocomplexes were visualized using an ECL substrate kit (Amersham International, Amersham, UK). Quantification of the target protein bands was through the ImageJ software measuring the total gray. The value was normalized to the loading control.
Statistical analyses
For statistical analyses the SPSS software (version 18.0) was employed. All values were expressed as the mean ± standard deviation (SD). All differences were considered significant at P < 0.05.
Author Contributions
All listed authors designed the study, performed the experiments and the statistical analysis, and wrote the manuscript. All authors have read the manuscript and approved the final version.
Conflicts of Interest
The authors declare that they have no conflicts of interests.
Funding
This project was supported by Shanghai Minhang Science and Technology Commission (2017MWDXK03), by Minhang District Talent Development Special Fund Project (2017), and Minhang District Subject Pilot Talent Project.
References
- 1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018; 68:7–30. https://doi.org/10.3322/caac.21442 [PubMed]
- 2. Choi KS, Suh M. Screening for gastric cancer: the usefulness of endoscopy. Clin Endosc. 2014; 47:490–96. https://doi.org/10.5946/ce.2014.47.6.490 [PubMed]
- 3. Hashim D, Boffetta P, La Vecchia C, Rota M, Bertuccio P, Malvezzi M, Negri E. The global decrease in cancer mortality: trends and disparities. Ann Oncol. 2016; 27:926–33. https://doi.org/10.1093/annonc/mdw027 [PubMed]
- 4. Shah MA. Gastrointestinal cancer: targeted therapies in gastric cancer-the dawn of a new era. Nat Rev Clin Oncol. 2014; 11:10–11. https://doi.org/10.1038/nrclinonc.2013.231 [PubMed]
- 5. Wadhwa R, Song S, Lee JS, Yao Y, Wei Q, Ajani JA. Gastric cancer-molecular and clinical dimensions. Nat Rev Clin Oncol. 2013; 10:643–55. https://doi.org/10.1038/nrclinonc.2013.170 [PubMed]
- 6. Doucet-Beaupré H, Ang SL, Lévesque M. Cell fate determination, neuronal maintenance and disease state: the emerging role of transcription factors Lmx1a and Lmx1b. FEBS Lett. 2015; 589:3727–38. https://doi.org/10.1016/j.febslet.2015.10.020 [PubMed]
- 7. Dong W, Feng L, Xie Y, Zhang H, Wu Y. Hypermethylation-mediated reduction of LMX1A expression in gastric cancer. Cancer Sci. 2011; 102:361–66. https://doi.org/10.1111/j.1349-7006.2010.01804.x [PubMed]
- 8. Chang CC, Huang RL, Wang HC, Liao YP, Yu MH, Lai HC. High methylation rate of LMX1A, NKX6-1, PAX1, PTPRR, SOX1, and ZNF582 genes in cervical adenocarcinoma. Int J Gynecol Cancer. 2014; 24:201–09. https://doi.org/10.1097/IGC.0000000000000054 [PubMed]
- 9. Zhang X, Qian Y, Li F, Bei S, Li M, Feng L. microRNA-9 selectively targets LMX1A to promote gastric cancer cell progression. Biochem Biophys Res Commun. 2018; 505:405–12. https://doi.org/10.1016/j.bbrc.2018.09.101 [PubMed]
- 10. Feng L, Xie Y, Zhao Z, Lian W. LMX1A inhibits metastasis of gastric cancer cells through negative regulation of β-catenin. Cell Biol Toxicol. 2016; 32:133–39. https://doi.org/10.1007/s10565-016-9326-0 [PubMed]
- 11. Chao TK, Yo YT, Liao YP, Wang YC, Su PH, Huang TS, Lai HC. LIM-homeobox transcription factor 1, alpha (LMX1A) inhibits tumourigenesis, epithelial-mesenchymal transition and stem-like properties of epithelial ovarian cancer. Gynecol Oncol. 2013; 128:475–82. https://doi.org/10.1016/j.ygyno.2012.12.018 [PubMed]
- 12. Su HY, Lai HC, Lin YW, Chou YC, Liu CY, Yu MH. An epigenetic marker panel for screening and prognostic prediction of ovarian cancer. Int J Cancer. 2009; 124:387–93. https://doi.org/10.1002/ijc.23957 [PubMed]
- 13. Lai HC, Lin YW, Huang TH, Yan P, Huang RL, Wang HC, Liu J, Chan MW, Chu TY, Sun CA, Chang CC, Yu MH. Identification of novel DNA methylation markers in cervical cancer. Int J Cancer. 2008; 123:161–67. https://doi.org/10.1002/ijc.23519 [PubMed]
- 14. Niu ZS, Niu XJ, Wang WH. Long non-coding RNAs in hepatocellular carcinoma: potential roles and clinical implications. World J Gastroenterol. 2017; 23:5860–74. https://doi.org/10.3748/wjg.v23.i32.5860 [PubMed]
- 15. Huo X, Han S, Wu G, Latchoumanin O, Zhou G, Hebbard L, George J, Qiao L. Dysregulated long noncoding RNAs (lncRNAs) in hepatocellular carcinoma: implications for tumorigenesis, disease progression, and liver cancer stem cells. Mol Cancer. 2017; 16:165. https://doi.org/10.1186/s12943-017-0734-4 [PubMed]
- 16. Yang X, Xie X, Xiao YF, Xie R, Hu CJ, Tang B, Li BS, Yang SM. The emergence of long non-coding RNAs in the tumorigenesis of hepatocellular carcinoma. Cancer Lett. 2015; 360:119–24. https://doi.org/10.1016/j.canlet.2015.02.035 [PubMed]
- 17. Hao NB, He YF, Li XQ, Wang K, Wang RL. The role of miRNA and lncRNA in gastric cancer. Oncotarget. 2017; 8:81572–82. https://doi.org/10.18632/oncotarget.19197 [PubMed]
- 18. Sun M, Nie FQ, Wang ZX, De W. Involvement of lncRNA dysregulation in gastric cancer. Histol Histopathol. 2016; 31:33–39. [PubMed]
- 19. Chen L, Dzakah EE, Shan G. Targetable long non-coding RNAs in cancer treatments. Cancer Lett. 2018; 418:119–24. https://doi.org/10.1016/j.canlet.2018.01.042 [PubMed]
- 20. Arun G, Diermeier SD, Spector DL. Therapeutic Targeting of Long Non-Coding RNAs in Cancer. Trends Mol Med. 2018; 24:257–77. https://doi.org/10.1016/j.molmed.2018.01.001 [PubMed]
- 21. Qu S, Yang X, Li X, Wang J, Gao Y, Shang R, Sun W, Dou K, Li H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015; 365:141–48. https://doi.org/10.1016/j.canlet.2015.06.003 [PubMed]
- 22. Cui C, Lu Z, Yang L, Gao Y, Liu W, Gu L, Yang C, Wilson J, Zhang Z, Xing B, Deng D, Sun ZS. Genome-wide identification of differential methylation between primary and recurrent hepatocellular carcinomas. Mol Carcinog. 2016; 55:1163–74. https://doi.org/10.1002/mc.22359 [PubMed]
- 23. Yang L, Zheng LY, Tian Y, Zhang ZQ, Dong WL, Wang XF, Zhang XY, Cao C. C6 ceramide dramatically enhances docetaxel-induced growth inhibition and apoptosis in cultured breast cancer cells: a mechanism study. Exp Cell Res. 2015; 332:47–59. https://doi.org/10.1016/j.yexcr.2014.12.017 [PubMed]
- 24. Wang SS, Lv Y, Xu XC, Zuo Y, Song Y, Wu GP, Lu PH, Zhang ZQ, Chen MB. Triptonide inhibits human nasopharyngeal carcinoma cell growth via disrupting Lnc-RNA THOR-IGF2BP1 signaling. Cancer Lett. 2019; 443:13–24. https://doi.org/10.1016/j.canlet.2018.11.028 [PubMed]
- 25. Chen MB, Liu YY, Xing ZY, Zhang ZQ, Jiang Q, Lu PH, Cao C. Itraconazole-Induced Inhibition on Human Esophageal Cancer Cell Growth Requires AMPK Activation. Mol Cancer Ther. 2018; 17:1229–39. https://doi.org/10.1158/1535-7163.MCT-17-1094 [PubMed]