



Introduction
Thyroid cancer is a common malignant tumor of the head and neck and endocrine system [1], and papillary thyroid carcinoma (PTC) is the most common type, accounting for 80% of all thyroid cancer cases [2]. PTC is more common in high iodine countries [3]. In recent years, the incidence of PTC has increased gradually, but the reasons are not clear [4]. Therefore, the exploration of the potential pathogenesis of PTC will be helpful for the early diagnosis and treatment of PTC as well as improving the prognosis of patients.
MicroRNA (miRNA) is the first to be found in non-coding RNA having a function, which contributes to post-transcriptional regulation of various genes by inhibiting the translation or degradation of mRNA [5]. Therefore, the dysregulation of miRNA is closely related to multiple diseases, including different types of cancer [6]. Changes in the expression of many miRNAs are associated with the ability of cancer to metastasize and invade [7]. Current studies have demonstrated that PTC is associated with overexpression of special miRNAs, such as miR-146b and miR-222 [8]. MiR-34a, a member of miR-34 family, was identified as a p53 target in 2007 [9], and extensive research has demonstrated miR-34a as an essential mediator of p53 functions and a potent tumor suppressor [10]. Recent study revealed that miR-34a was downregulated in human pancreatic cancer, and miR-34a might act as a biomarker for the early diagnosis of pancreatic cancer [11]. Furthermore, Liu found that miR-34a was downregulated in thyroid cancer, XIST regulated the expression of miR-34a through the competitive endogenous RNAs (ceRNA) mechanism, and ultimately regulates the proliferation of thyroid cancer cells and tumor growth [12]. Nevertheless, miR-34a’s function in PTC has not been validated, and the underlying mechanism in PTC remains unclear.
Compared miRNAs, our understanding of long noncoding RNAs (lncRNAs) is still in its infancy, and little is known about the diagnostic or prognostic value of lncRNAs in PTC. Interestingly, the cross-regulatory network between miRNAs and lncRNAs has recently been discovered, which provides insights into the function of these non-coding RNAs and their potential as therapeutic approaches [13, 14]. Recent studies have reported the dysregulated expression of several lncRNAs in PTC. For example, PVT1 regulates the proliferation of thyroid cancer cells by recruiting EZH2 and TSHR [15]. Therefore, we aimed to further explore the upstream molecular mechanism by which miR-34a regulates PTC, and whether lncRNAs are involved in the progress of PTC by regulating miR-34a activity.
In this study, we found that miR-34a directly targeted SOX4, thereby inhibiting the development of PTC. Furthermore, we first identified the lncRNA FOXN3-AS2, and found that upregulation of FOXN3-AS2 promoted the growth of PTC via miR-34a/SOX4 axis.
Results
Expression of miR-34a in PTC tissues and cell lines
To evaluate the effects of miR-34a in PTC, we firstly detected miR-34a expression in PTC clinical tissues and cell lines. As shown in Figure 1A, the mRNA level of miR-34a was decreased in PTC tissues comparing with adjacent normal tissues. In addition, we examined miR-34a expression in PTC cell lines. Compared with normal thyroid gland cells Nthy-ori 3-1, miR-34a expressions in PTC cell lines HIM-1 and TCP-1 were remarkably reduced (Figure 1B). Then, we analyzed the 60-months survival rate of PTC patients. According to the average expression of miR-34a in 1A, the 40 PTC patients were divided into low and high miR-34a expression groups. The patients with low miR-34a expression had a lower survival rate (Figure 1C). These data suggested a potential regulatory role of miR-34a in the pathological process of PTC.
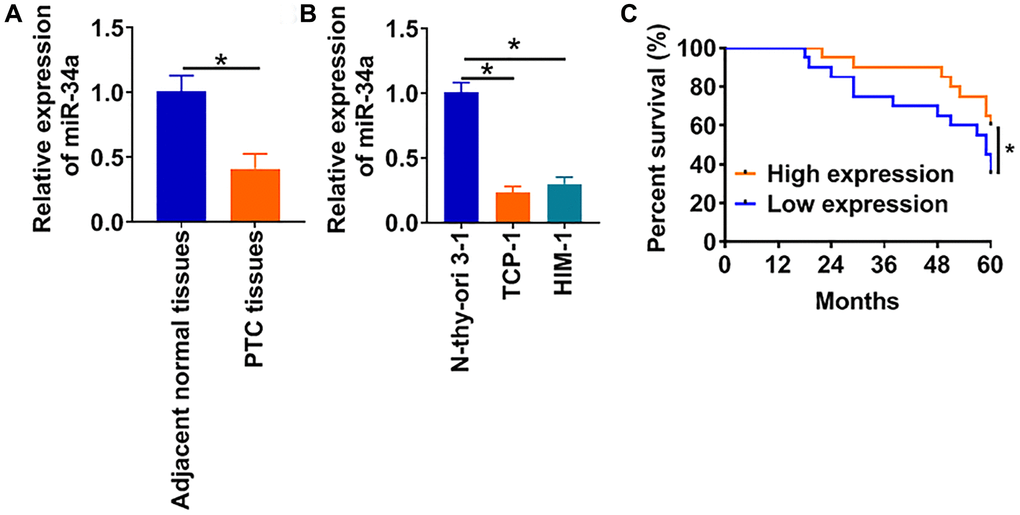
Figure 1. LncRNA FGD5-AS1 is increased in OC tissue and cells. (A) The expression of miR-34a in PTC tissues (n = 40) and adjacent normal tissues (n = 40) determined by qRT-PCR (*p < 0.05). (B) qRT-PCR assay analyzed the expression of miR-34a in normal human thyroid cells (Nthy-ori 3-1) and PTC cell lines (TCP-1 and HIM-1) (*p < 0.05). (C) The 60-months survival rate of PTC patients with high and low miR-34a expression (*p < 0.05).
Overexpression of miR-34a suppresses proliferation, migration and invasion of PTC cells
To further explore the role of miR-34a in PTC, miR-34a mimics or its negative control (NC) was transfected into TCP-1 cells. The transfection of miR-34a mimics significantly increased miR-34a expression (Figure 2A). Next, we used CCK8 and colony formation assay to determine cell proliferation. The overexpression of miR-34a inhibited the proliferated capacity of TCP-1 cells (Figure 2B and 2C). Migration and invasion are critical to the growth of cancer cells, and we then detected the migration and invasion of TCP-1 cells upon miR-34a mimics or NC transfection. The wound healing results showed miR-34a significantly suppressed cell migration (Figure 2D). Meanwhile, transwell assay uncovered a lower invasive ability in miR-34a mimics transfected TCP-1 cells (Figure 2E).
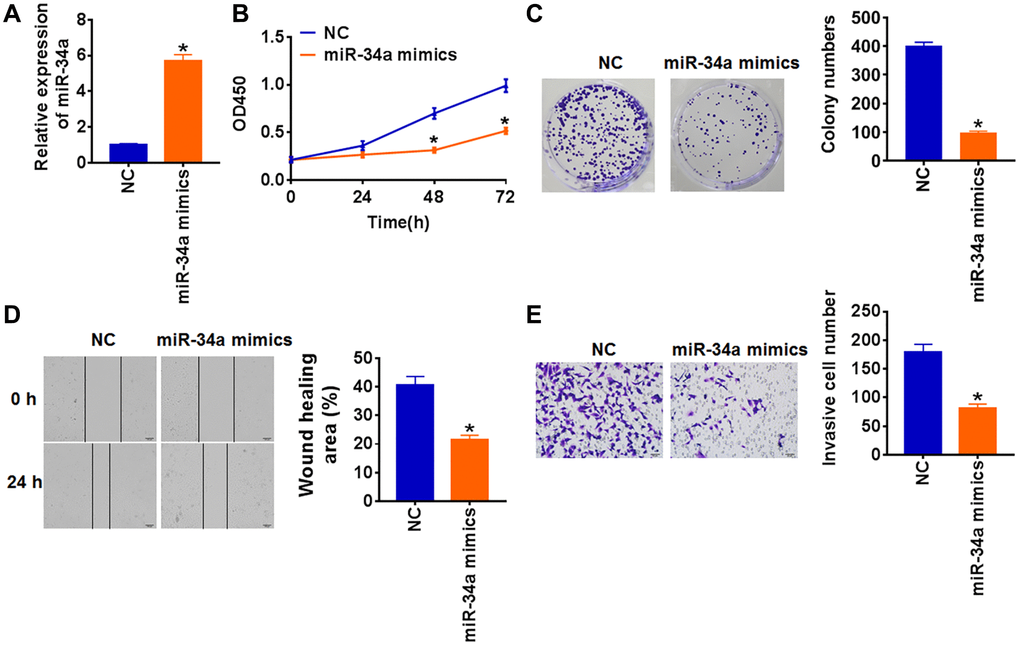
Figure 2. Upregulation of miR-34a inhibits proliferation, migration and invasion in TCP-1 cells. miR-34a mimics and its negative control (NC) was transfected into TCP-1 cells. (A) The expression of miR-34a was determined by qRT-PCR (*p < 0.05). (B) CKK-8 assay was used to examine the cell growth at 0, 24, 48 and 72 h (*p < 0.05). (C) Colony formation assay was used to calculated cell proliferation (*p < 0.05). (D) Wound healing assay was used to detect cell migration (*p < 0.05). (E) Transwell assay was performed to check cell invasive ability (*p < 0.05).
MiR-34a suppresses PTC progression by targeting SOX4 in PTC cells
SOX4 is abnormally expressed in a variety of tumor tissues, and is closely related to the occurrence, development and metastasis of tumors. In our study, we surprisedly found that there were complementary base pairs between SOX4 and miR-34a using multiple databases (Targetscan, miRanda and TarBase) (Figure 3A), which revealed SOX4 might be a potential target of miR-34a in regulating PTC progression. To test our hypothesis, we performed luciferase assay and found that miR-34a mimics significantly decreased luciferase activity of WT 3′UTR of SOX4 rather than mutant SOX4 (Figure 3B). Furthermore, miR-34a mimics inhibited both mRNA and protein level of SOX4, while miR-34a inhibitor increased SOX4 expression (Figure 3C and 3D). Moreover, we found an obvious increase of SOX4 protein in PTC tissues and cells (Figure 3E and 3F). Besides, we analyzed the correlation of miR-34a and SOX4 expression in PTC tissues, which showed that miR-34a expression was negatively related to SOX4 level (Figure 3G).
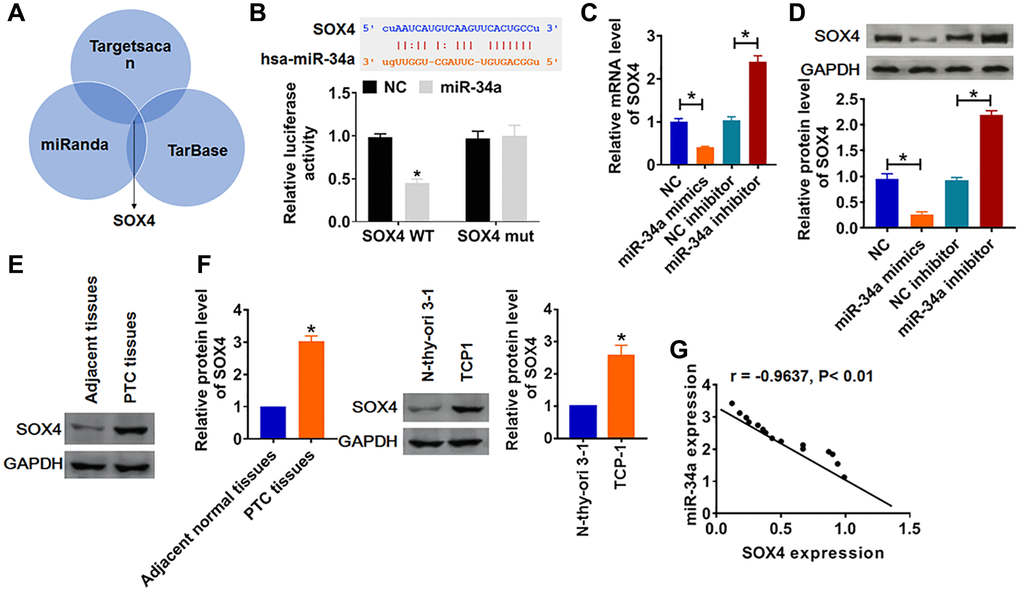
Figure 3. SOX4 was a direct target of miR-34a. (A) Multiple databases (Targetscan, miRanda and TarBase) identified that the SOX4 had complementary base pairs with miR-34a. (B) WT and mutant SOX4 luciferase plasmids were transfected into HEK293 cells with miR-34a mimics or NC. The luciferase activity was measured by dual-luciferase reporter assay system. (*p < 0.05). miR-34a or miR-34a inhibitor or its NC was transfected into TCP-1 cells. (C) The mRNA level of SOX4 was analyzed by qRT-PCR, (D) and the protein level of SOX4 was tested by Western blot (*p < 0.05). The protein expression of SOX4 in PTC tissues (E) and cells (F) was detected by Western blot (*p < 0.05). (G) The correlation analysis of miR-34a and SOX4 expression in PTC tissues.
To verify whether miR-34a regulated PTC by targeting SOX4, we forced expression of SOX4 in TCP-1 cell (Figure 4A and 4B), and cotransfected miR-34a mimics. As shown in Figure 4C and 4D, upregulation of SOX4 reversed the inhibitory effect of miR-34a on TCP-1 cells’ proliferation. SOX4 also restored the migrative and invasive ability (Figure 4E and 4F). These data indicated that miR-34a inhibited the malignancy of PTC cells via inhibiting SOX4.
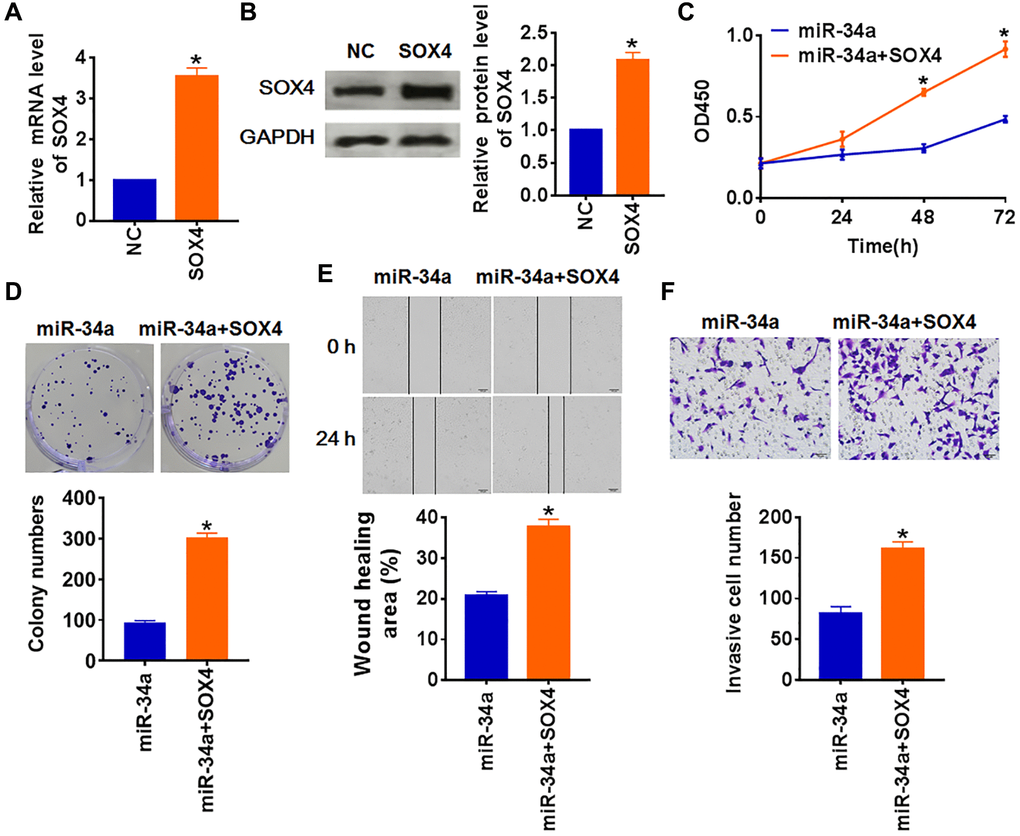
Figure 4. MiR-34a inhibits PTC development by downregulating SOX4. SOX4 or NC was transfected into TCP-1 cells. (A) The mRNA expression of SOX4 was determined by qRT-PCR, (B) and the protein level of SOX4 was tested by Western blot(*p < 0.05). (C) CKK-8 assay was used to examine the cell growth at 0, 24, 48 and 72 h (*p < 0.05). (D) Colony formation assay was used to calculated cell proliferation (*p < 0.05). (E) Wound healing assay was used to detect cell migration (*p < 0.05). (F) Transwell assay was performed to check cell invasive ability (*p < 0.05).
FOXN3-AS2 acts as a sponge of miR-34a
Evidence is mounting that lncRNAs competitively bind to miRNAs and further inhibit its activity [16]. Thus, we used Starbase, LncBook and DIANA databases to identify the lncRNAs with miR-34a biding sites. Our data revealed that lncRNA FOXN3-AS2 could bind with miR-34a (Figure 5A). Following luciferase assay showed that overexpression of miR-34a increased luciferase activity of WT FOXN3-AS2 but not Mut one (Figure 5B). In addition, FOXN3-AS2 or si-FOXN3-AS2 was transfected into TCP-1 cells. FOXN3-AS2 plasmid promoted FOXN3-AS2 level, and si-FOXN3-AS2 inhibited FOXN3-AS2 expression (Figure 5C). Meanwhile, FOXN3-AS2 suppressed miR-34a and increased SOX4 expression, while silencing of FOXN3-AS2 had the opposite effect (Figure 5C). Then, FOXN3-AS2 plasmid and miR-34a were cotransfected into TCP-1 cells, which showed that miR-34a abolished the promoting role of FOXN3-AS2 on SOX4 expression (Figure 5D). These data suggested FOXN3-AS2 promoted SOX4 expression via inhibiting miR-34a. What’s more, FOXN3-AS2 was highly expressed in PTC tissues and cells (Figure 5E and 5F). In addition, the expression of FOXN3-AS2 expression in PTC tissues was negatively related to miR-34a expression (Figure 5G). Together, FOXN3-AS2 promoted SOX4 expression via inhibiting miR-34a.
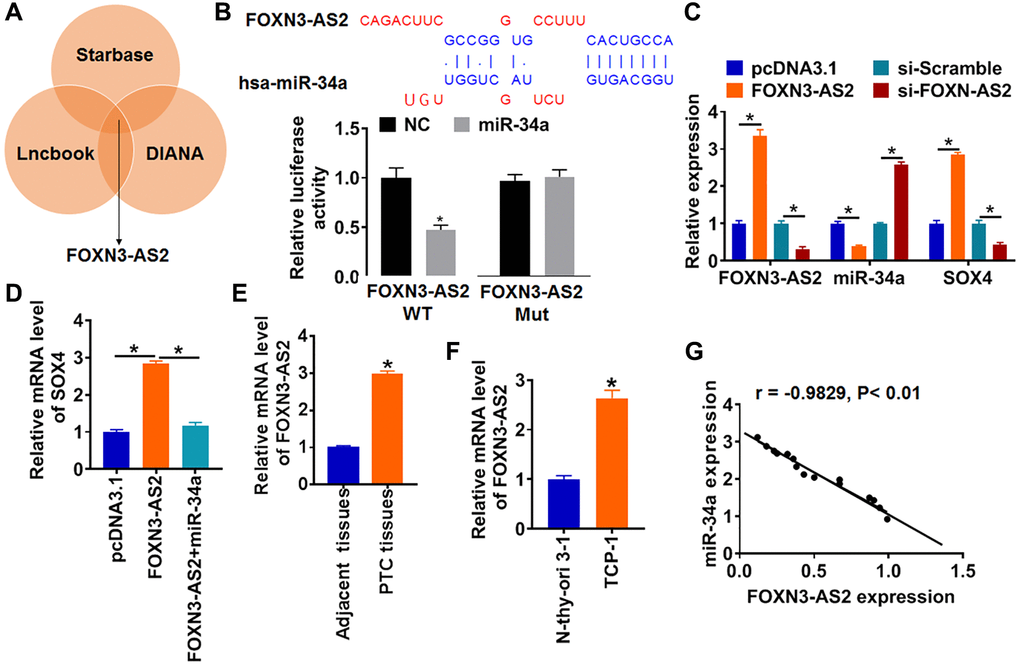
Figure 5. FOXN3-AS2 directly regulates miR-34a and inhibits its expression in PTC. (A) Starbase, LncBook and DIANA databases used to identify the lncRNAs with miR-34a biding sites. (B) WT and mutant miR-34a luciferase plasmids were transfected into HEK293 cells with pcDNA3.1 or FOXN3-AS2. The luciferase activity was measured by dual-luciferase reporter assay system (*p < 0.05). (C) FOXN3-AS2 or si-FOXN3-AS2 or its NC was transfected into TCP-1 cells. The mRNA level of FOXN3-AS2, miR-34a and SOX4 were analyzed by qRT-PCR (*p < 0.05). (D) FOXN3-AS2 and miR-34a were cotransfected into TCP-1 cells, the mRNA level of SOX4 were tested (*p < 0.05). The protein level of miR-34a in PTC tissues (E) and cells (F) was detected by qRT-PCR (*p < 0.05). (G) The correlation analysis of miR-34a and FOXN3-AS2 expression in PTC tissues.
FOXN3-AS2 promotes proliferation, migration and invasion via miR-34a/SOX4 axis in PTC cells
To assess the role of FOXN3-AS2, we transfected FOXN3-AS2 plasmid into TCP-1 cells, and qRT-PCR assay showed an increase of FOXN3-AS2 level (Figure 6A). Next, we cotransfected miR-34a mimics or si-SOX4 with FOXN3-AS2 plasmid into TCP-1 cells. Followed functional experiments showed FOXN3-AS2 promoted proliferation, migration and invasion of TCP-1 cells, while this facilitated effect was removed by miR-34a mimics or si-SOX4 (Figure 6B–6E). Taken together, FOXN3-AS2 regulated the progression of PTC via miR-34a/SOX4 axis.
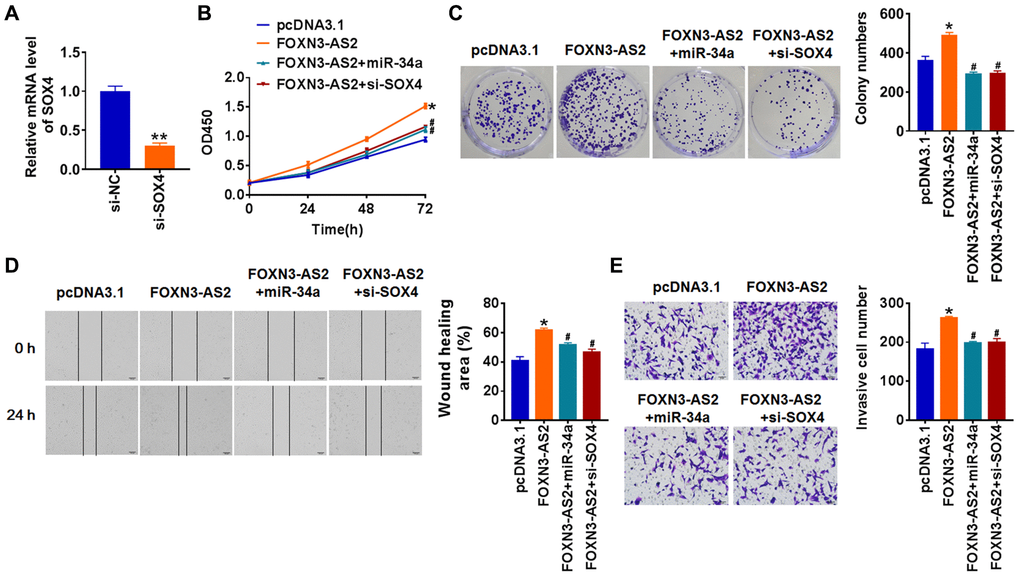
Figure 6. LncRNA FOXN3-AS2 promotes OC progression via miR-34a/SOX4 axis. (A) FOXN3-AS2 plasmid was transfected into TCP-1 cells, and qRT-PCR analysis was used to detect the transfection efficiency (*p < 0.05). miR-34a or si-SOX4 was cotransfected into TCP-1 cells with FOXN3-AS2 plasmid. CCK8 assay (B) and colony formation assay (C) was used to calculate cell proliferation (*p < 0.05 vs. pcDNA3.1, #p < 0.05 vs. FOXN3-AS2). (D) Wound healing assay was used to detect cell migration (*p < 0.05 vs. pcDNA3.1, #p < 0.05 vs. FOXN3-AS2). (E) Transwell assay was performed to check cell invasive ability (*p < 0.05 vs. pcDNA3.1, #p < 0.05 vs. FOXN3-AS2).
FOXN3-AS2 promotes in vivo tumor growth in the nude mice
For further explore the function of FOXN3-AS2 in PTC tumor, we set up xenograft nude mice model. 30 mice were divided into two group randomly, TCP-1 cells stably expressing FOXN3-AS2 or pcDNA3.1 were subcutaneously injected in right lower limb of the nude mice. The mice with FOXN3-AS2 transfected cells showed a bigger tumor volume, and tumors grew faster (Figure 7A). The tumors were isolated at 30 days after injection, FOXN3-AS2 significantly increased tumors weight than that pcDNA3.1 (Figure 7B). In addition, we isolated these tumor tissues and found that the expression of FOXN3-AS2 was increased in FOXN3-AS2 overexpression mice (Figure 7C). Moreover, FOXN3-AS2 decreased the mRNA level miR-34a, but increased SOX4 expression in tumor tissues (Figure 7D and 7E). These data indicated that FOXN3-AS2 accelerated PTC tumorigenesis via modulating miR-34a/SOX4 axis.
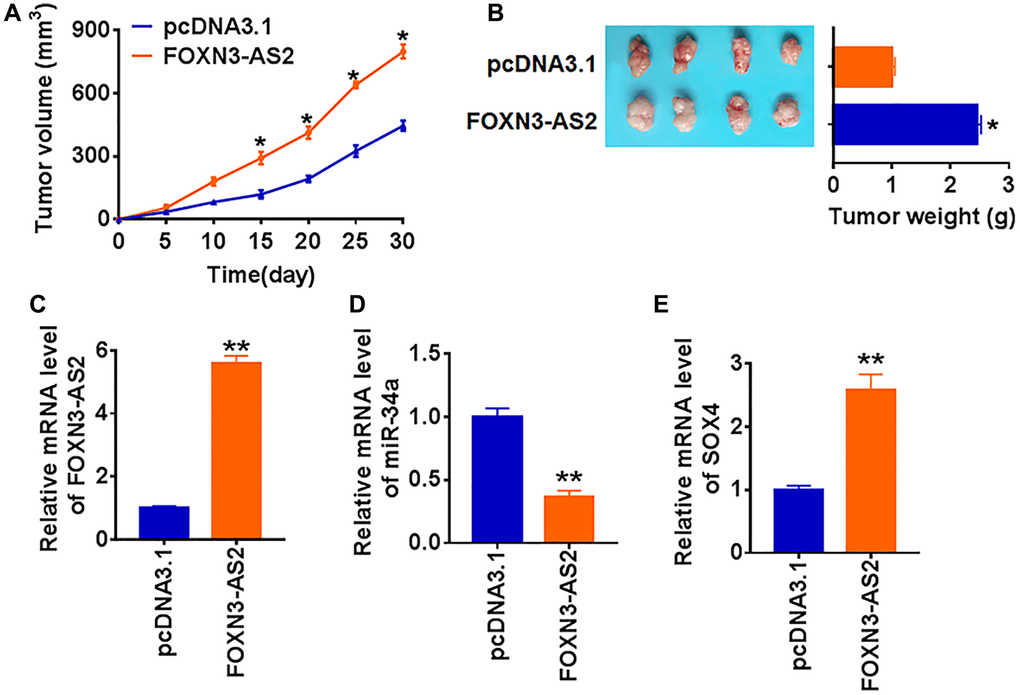
Figure 7. LncRNA FOXN3-AS2 promotes in vivo tumor growth in the nude mice. TCP-1 cells line stably expressing FOXN3-AS2 or pcDNA3.1 were subcutaneously injected in right lower limb of the nude mice. (A) The tumor volume was assessed in the nude mice every 5 days (*p < 0.05). (B) Tumor weight was determined in the isolated tumors from the nude mice (*p < 0.05). (C) The relative expression of FOXN3-AS2 was determined by qRT-PCR in the isolated tumor tissues (**p < 0.01). qRT-PCR was performed to detect the relative mRNA expression of miR-34a (D) and SOX4 (E) (*p < 0.05).
Discussion
Among all types of thyroid cancer, PTC accounts for the highest proportion [17]. Although the prognosis of early PTC patients is satisfactory, the five-year survival rate of late PTC patients is low [18]. Therefore, it is imperative to explore the underlying mechanism of PTC and identify more effective therapeutic strategies.
MiRNAs are widely found in eukaryotic cells and perform different biological functions by targeting and binding different miRNAs [19]. Previous studies have shown that miRNAs play an important role in the occurrence and development of various malignant tumors [20]. For example, miR-146b was significantly increased in PTC. MiR-146b induced TPC-1 cells to enter S phase, and the cells were blocked in the G0 phase, thus promoting the proliferation of cancer cells [21]. Being a member of the miR-34 family, miR-34a was identified as a p53 target and acted as tumor suppressor in multiple cancers [22]. Present study focused on the role of miR-34a in PTC progression. miR-34a was significantly lower expressed in PTC tissues than that in normal thyroid tissues. Meanwhile, miR-34a was increased in PTC cell line. Differential expression of miR-34a indicated a potential of miR-34a in regulating the development of PTC. Thus, we forced expression of miR-34a in TPC-1 cells, and found upregulation of miR-34a inhibited cell proliferation, migration and invasion. Our finding revealed miR-34a acted as an anti-tumor gene, which was consistent with the role of miR-34a in pancreatic ductal adenocarcinoma and Pituitary adenomas [23, 24]. However, miR-34a has also been reported to be up-regulated in PTC tissues and cell lines [25–27]. Bonetti et al. found that miR-34a had a dual role in the control of early progenitor proliferation and commitment in the mammary gland and in breast cancer. The separate functions of miR-34a might result from the multiple upstream regulators [28]. Thus, we speculated that the expression of miR-34a in PTC were also regulated by multiple upstream regulators, some factors promoted miR-34a level, while others inhibited miR-34a expression. The final expression of miR-34a was determined by the level of the positive or negative upstream regulators. Factors such as underlying diseases and living conditions may influence the level of upstream regulators in PTC patients, which then induced the opposite expression of miR-34a.
Studies have shown that miRNAs are involved in the regulation of various diseases through targeted inhibition of mRNA expression. To illustrate the mechanism of miR-34a in PTC progression, the commonly used databases predicted the potential target of miR-34a. Interestingly, we found SOX4 was a direct target of miR-34a, and miR-34a inhibited SOX4 expression in TPC-1 cells. SOX4 is a family member of SOX transcription factors [29]. SOX4 takes part in the progress of embryo development, development of cardiovascular and immune system [30–32]. In addition, recent studies show that SOX4 improves tumor growth [33]. Present study showed a remarkable elevation of SOX4 in PTC tissues and cells. Meanwhile, overexpression of SOX4 removed the inhibitory effect of miR-34a on PTC. These data indicated that miR-34a acted as a tumor suppressor of PTC by inhibiting SOX4 expression.
LncRNAs are proved to function as a sponge of miRNAs and inhibited their expression and activity in cancer development, and lncRNAs act as the upstream regulators [34, 35]. Thus, we searched for a lncRNA sponged miR-34a to fully uncovered the mechanism of miR-34a modulating PTC. LncRNA FOXN3-AS2 was expression by inhibiting miR-34a expression. FOXN3-AS2 is the antisense of FOXN3, FOXN3 is involved in the progression of PTC by inactivating Wnt/β-catenin pathway [36]. However, there is little study about FOXN3-AS2 in tumor growth. Present study first found that FOXN3-AS2 was increased in PTC tissues and cells, and FOXN3-AS2 promoted SOX4 expression by sponging miR-34a. The following functional experiments indicated that FOXN3-AS2 promoted PTC progression both in vitro and in vivo, and the underlying mechanism might be regulating miR-34a/SOX4 axis.
At present, the role and mechanism of ncRNAs are gradually clarified. Differential expression of ncRNAs may act as potential marker in differentiating thyroid cancer types, stages and metastases. With the development of molecular biology, people will have a deeper understanding of ncRNAs, which will benefit the diagnosis, treatment and prognosis of PTC.
Conclusions
In conclusion, our results clarified the lncRNA FOXN3-AS21 function for the first time, and FOXN3-AS2 promoted the progression of PTC, which was mediated by miR-34a/SOX4 axis. This study might provide new understanding and targets for FOXN3-AS2 diagnosis and treatment.
Materials and Methods
Clinical samples
Fresh cancer tissue samples and adjacent normal tissue samples were taken from 40 PTC patients undergoing surgical procedures at Jilin Cancer Hospital. All patients did not receive radiotherapy, chemotherapy, and immunization before cancer tissue collection, and all tissues were confirmed by pathological examination. All of the patients or their guardians provided written consent, and our experiments were supported by the Ethics Committee and Institutional Review Board of Jilin Cancer Hospital.
Cell culture
The cell lines (Nthy-ori 3-1, HIM-1 and TCP-1) were purchased from the Science Cell Laboratory. Cells were cultured in RPMI 1640 (GIBCO, USA) supplemented with 10% fetal bovine serum (Cromwell, USA) and 100 μL/mL penicillin and streptomycin (Sigma-Aldrich, USA) and placed at 37°C with 5% CO2.
Cell transfection
About 5 × 105 cells per well were seeded in 6-well plates, 2000 ng small interference RNA (siRNA) or negative control of siRNA (si-NC), 2000 ng plasmid or empty vector (pcDNA3.1), 20 nmol miRNA mimics or negative control (NC) of miRNA mimics, and 20 nmol miRNA inhibitor or negative control of miRNA inhibitor (NC inhibitor) were transfected into the cells for 48 h using Lipo2000 (Thermo Fisher Scientific, USA) according to the manufacturer’s recommendations.
qRT-PCR
The RNA Isolation Kit (Vazyme, China) was used to extract total RNA from tissues or cells, and the concentration and quality of the purified RNA were measured using Nanodrop 2000/2000C spectrophotometer. RNA was reverse transcribed into cDNA template at the following reaction temperatures: 37°C 15 min, 50°C 5 min, 98°C 5 min, 4°C hold. The cDNA was stored at −20°C.Then the primers synthesized by Shanghai Jikegein Company were used for quantitative PCR. The reaction system of 20 μL was prepared. The reaction was carried out according to the following steps: pre-denaturation at 95°C for 15 min, denaturation at 95°C for 15s and annealing extension at 60°C for 1min for 35–40 cycles. LncRNA FOXN3-AS2 and SOX4 were standardized by GAPDH, and miR-34a was standardized by U6. The expression l of FOXN3-AS2 and SOX4 and miR-34a was analyzed by 2−ΔΔCT.
Western blot
The tissues or cells in each group were collected and treated with RIPA lysate. Then, the protein concentration was evaluated by BCA kit (Beyotime, China). And the proteins in loading buffer were separated by SDS-PAGE and transferred to PVDF membrane. The membrane carrying proteins was sealed with blocking fluid, and was incubated with SOX4 (1: 500) and GAPDH (1: 1000) overnight at 4°C. The next day, HRP-labeled secondary antibody (1: 5000) was added and incubated at room temperature for 2 h. The membrane was photographed by ECL exposure and analyzed the gray value of the target bands. The primary antibodies were purchased from Abcam, SOX4 (ab80261), and GAPDH (ab8245) was used as an internal control.
CCK8 assay
The logarithmic growth phase cells were digested by trypsin and resuspended in complete medium and counted. Cells were inoculated in a 96-well plate at a density of 2500 cells/well and cultured in a cell incubator. From the second day to the fifth day after planking, 10 μl CCK-8 reagent (Vazyme, China) was added 2–4 h before culture termination every day. After 4 h, the 96-well plate was placed on the oscillator and oscillated for 5 min. The OD value was detected at 450 nm with a microplate analyzer. The final values of each group were plotted and the results were analyzed.
Wound healing assay
5 × 105 cells were planted in a 6-well plate, and when the cells grew to fuse, two vertical parallel lines were drawn with 10 μL suction heads against the ruler. The floating cells were washed with PBS and cultured in serum-free medium for 24 h. Images were taken at 0 and 24 h of cell culture, respectively.
Transwell assay
Cells in logarithmic growth phase were adjusted to 2 × 105 cells/well of medium (without serum) and plated into the upper chamber insert pre-coated with 1 μg/μl Matrigel. Lower chamber was added with 500 μL of medium (with 10% FBS), and then incubate the chamber at 37°C for 48 h. Then the invading cells were visualized by the crystal violet and inverted microscope.
Colony formation assay
Cells were seeded into 6-well plates at a density of 100 cells/well and cultured for 14 days. Cells clone were fixed with 4% paraformaldehyde and stained with 0.1% Crystal Violet (Sigma-Aldrich. MO, USA) at room temperature for 15 min. Then cells were rinsed with distilled water, and the colonies were visualized by inverted microscope.
Dual luciferase reporter assay
psiCHECK-2 luciferase reporter plasmid was inserted with the wildtype (WT) 3′UTR of SOX4/FOXN3-AS2 or mutant (Mut) 3′UTR SOX4/FOXN3-AS2 sequences that contain the putative binding sites of miR-34a. NC or miR-34a mimics were transfected with reporter vectors into HEK293 cells. The cells were collected after 48 h post-transfection and lysed to detect the luciferase activity (Promega).
In vivo tumor growth assay
Nude mice were purchased from Beijing Charles river. The TCP-1 cell line stably expressing FOXN3-AS2 or pcDNA3.1 were constructed by Cyagen Biosciences (China). The TCP-1 cells (5 × 106) were subcutaneously injected in right lower limb of the nude mice. Tumor size was measured every five days. After 30 days of cells injection, mice were killed and the tumors were removed for follow-up study. The animal experimental procedures were approved by the animal experimental ethics committee of Jilin Cancer Hospital, and the experiments were carried out following the Guide for the Care and Use of Experimental Animals.
Statistical analysis
Significant differences were calculated using two-tailed t-test through Graphpad 8.0. The P values less than 0.05 were considered significance.
Author Contributions
Xiao-Song Sun and Zheng-Chao Wei designed and performed experiments, analysed data and wrote the paper; Shuang Liu and Lei Shi designed and performed experiments; Yan-Peng Li and Ming Wang designed experiments, analysed data and wrote the paper.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
- 1. Anania MC, Di Marco T, Mazzoni M, Greco A. Targeting Non-Oncogene Addiction: Focus on Thyroid Cancer. Cancers (Basel). 2020; 12:129. https://doi.org/10.3390/cancers12010129 [PubMed]
- 2. Lewiński A, Adamczewski Z, Zygmunt A, Markuszewski L, Karbownik-Lewińska M, Stasiak M. Correlations between Molecular Landscape and Sonographic Image of Different Variants of Papillary Thyroid Carcinoma. J Clin Med. 2019; 8:1916. https://doi.org/10.3390/jcm8111916 [PubMed]
- 3. Kiriakopoulos A, Petralias A, Linos D. Multifocal Versus Solitary Papillary Thyroid Carcinoma. World J Surg. 2016; 40:2139–43. https://doi.org/10.1007/s00268-016-3628-5 [PubMed]
- 4. Sohn SY, Choi JY, Jang HW, Kim HJ, Jin SM, Kim SW, Suh S, Hur KY, Kim JH, Chung JH, Kim SW. Association between excessive urinary iodine excretion and failure of radioactive iodine thyroid ablation in patients with papillary thyroid cancer. Thyroid. 2013; 23:741–47. https://doi.org/10.1089/thy.2012.0136 [PubMed]
- 5. Cheng M, Wang B, Yang M, Ma J, Ye Z, Xie L, Zhou M, Chen W. microRNAs expression in relation to particulate matter exposure: A systematic review. Environ Pollut. 2020; 260:113961. https://doi.org/10.1016/j.envpol.2020.113961 [PubMed]
- 6. Yu X, Li Z, Shen J, Chan MT, Wu WK. Role of microRNAs in primary central nervous system lymphomas. Cell Prolif. 2016; 49:147–53. https://doi.org/10.1111/cpr.12243 [PubMed]
- 7. Li X, Han J, Zhu H, Peng L, Chen Z. miR-181b-5p mediates TGF-β1-induced epithelial-to-mesenchymal transition in non-small cell lung cancer stem-like cells derived from lung adenocarcinoma A549 cells. Int J Oncol. 2017; 51:158–68. https://doi.org/10.3892/ijo.2017.4007 [PubMed]
- 8. Benvenga S, Koch CA. Molecular pathways associated with aggressiveness of papillary thyroid cancer. Curr Genomics. 2014; 15:162–70. https://doi.org/10.2174/1389202915999140404100958 [PubMed]
- 9. Raver-Shapira N, Marciano E, Meiri E, Spector Y, Rosenfeld N, Moskovits N, Bentwich Z, Oren M. Transcriptional activation of miR-34a contributes to p53-mediated apoptosis. Mol Cell. 2007; 26:731–43. https://doi.org/10.1016/j.molcel.2007.05.017 [PubMed]
- 10. Li WJ, Wang Y, Liu R, Kasinski AL, Shen H, Slack FJ, Tang DG. MicroRNA-34a: Potent Tumor Suppressor, Cancer Stem Cell Inhibitor, and Potential Anticancer Therapeutic. Front Cell Dev Biol. 2021; 9:640587. https://doi.org/10.3389/fcell.2021.640587 [PubMed]
- 11. Akula SM, Ruvolo PP, McCubrey JA. TP53/miR-34a-associated signaling targets SERPINE1 expression in human pancreatic cancer. Aging (Albany NY). 2020; 12:2777–97. https://doi.org/10.18632/aging.102776 [PubMed]
- 12. Liu H, Deng H, Zhao Y, Li C, Liang Y. LncRNA XIST/miR-34a axis modulates the cell proliferation and tumor growth of thyroid cancer through MET-PI3K-AKT signaling. J Exp Clin Cancer Res. 2018; 37:279. https://doi.org/10.1186/s13046-018-0950-9 [PubMed]
- 13. Liang H, Su X, Wu Q, Shan H, Lv L, Yu T, Zhao X, Sun J, Yang R, Zhang L, Yan H, Zhou Y, Li X, et al. LncRNA 2810403D21Rik/Mirf promotes ischemic myocardial injury by regulating autophagy through targeting Mir26a. Autophagy. 2020; 16:1077–91. https://doi.org/10.1080/15548627.2019.1659610 [PubMed]
- 14. Schulte C, Barwari T, Joshi A, Theofilatos K, Zampetaki A, Barallobre-Barreiro J, Singh B, Sörensen NA, Neumann JT, Zeller T, Westermann D, Blankenberg S, Marber M, et al. Comparative Analysis of Circulating Noncoding RNAs Versus Protein Biomarkers in the Detection of Myocardial Injury. Circ Res. 2019; 125:328–40. https://doi.org/10.1161/CIRCRESAHA.119.314937 [PubMed]
- 15. Zhou Q, Chen J, Feng J, Wang J. Long noncoding RNA PVT1 modulates thyroid cancer cell proliferation by recruiting EZH2 and regulating thyroid-stimulating hormone receptor (TSHR). Tumour Biol. 2016; 37:3105–13. https://doi.org/10.1007/s13277-015-4149-9 [PubMed]
- 16. Wang Y, Yang L, Chen T, Liu X, Guo Y, Zhu Q, Tong X, Yang W, Xu Q, Huang D, Tu K. A novel lncRNA MCM3AP-AS1 promotes the growth of hepatocellular carcinoma by targeting miR-194-5p/FOXA1 axis. Mol Cancer. 2019; 18:28. https://doi.org/10.1186/s12943-019-0957-7 [PubMed]
- 17. He J, Zhou M, Li X, Gu S, Cao Y, Xing T, Chen W, Chu C, Gu F, Zhou J, Jin Y, Ma J, Ma D, Zou Q. SLC34A2 simultaneously promotes papillary thyroid carcinoma growth and invasion through distinct mechanisms. Oncogene. 2020; 39:2658–75. https://doi.org/10.1038/s41388-020-1181-z [PubMed]
- 18. Nixon IJ, Wang LY, Ganly I, Patel SG, Morris LG, Migliacci JC, Tuttle RM, Shah JP, Shaha AR. Outcomes for patients with papillary thyroid cancer who do not undergo prophylactic central neck dissection. Br J Surg. 2016; 103:218–25. https://doi.org/10.1002/bjs.10036 [PubMed]
- 19. Dong Y, Xiao Y, Shi Q, Jiang C. Dysregulated lncRNA-miRNA-mRNA Network Reveals Patient Survival-Associated Modules and RNA Binding Proteins in Invasive Breast Carcinoma. Front Genet. 2020; 10:1284. https://doi.org/10.3389/fgene.2019.01284 [PubMed]
- 20. Duran-Sanchon S, Moreno L, Gómez-Matas J, Augé JM, Serra-Burriel M, Cuatrecasas M, Moreira L, Serradesanferm A, Pozo À, Grau J, Pellisé M, Gironella M, Castells A. Fecal MicroRNA-Based Algorithm Increases Effectiveness of Fecal Immunochemical Test-Based Screening for Colorectal Cancer. Clin Gastroenterol Hepatol. 2021; 19:323–30.e1. https://doi.org/10.1016/j.cgh.2020.02.043 [PubMed]
- 21. Wojtas B, Ferraz C, Stokowy T, Hauptmann S, Lange D, Dralle H, Musholt T, Jarzab B, Paschke R, Eszlinger M. Differential miRNA expression defines migration and reduced apoptosis in follicular thyroid carcinomas. Mol Cell Endocrinol. 2014; 388:1–9. https://doi.org/10.1016/j.mce.2014.02.011 [PubMed]
- 22. Chen S, Huang A, Chen H, Yang Y, Xia F, Jin L, Zhang J. miR-34a inhibits the apoptosis of MDSCs by suppressing the expression of N-myc. Immunol Cell Biol. 2016; 94:563–72. https://doi.org/10.1038/icb.2016.11 [PubMed]
- 23. Chen Y, Li Z, Zhang M, Wang B, Ye J, Zhang Y, Tang D, Ma D, Jin W, Li X, Wang S. Circ-ASH2L promotes tumor progression by sponging miR-34a to regulate Notch1 in pancreatic ductal adenocarcinoma. J Exp Clin Cancer Res. 2019; 38:466. https://doi.org/10.1186/s13046-019-1436-0 [PubMed]
- 24. Yang Z, Zhang T, Wang Q, Gao H. Overexpression of microRNA-34a Attenuates Proliferation and Induces Apoptosis in Pituitary Adenoma Cells via SOX7. Mol Ther Oncolytics. 2018; 10:40–47. https://doi.org/10.1016/j.omto.2018.07.001 [PubMed]
- 25. Ma Y, Qin H, Cui Y. MiR-34a targets GAS1 to promote cell proliferation and inhibit apoptosis in papillary thyroid carcinoma via PI3K/Akt/Bad pathway. Biochem Biophys Res Commun. 2013; 441:958–63. https://doi.org/10.1016/j.bbrc.2013.11.010 [PubMed]
- 26. Sheu SY, Grabellus F, Schwertheim S, Worm K, Broecker-Preuss M, Schmid KW. Differential miRNA expression profiles in variants of papillary thyroid carcinoma and encapsulated follicular thyroid tumours. Br J Cancer. 2010; 102:376–82. https://doi.org/10.1038/sj.bjc.6605493 [PubMed]
- 27. Cahill S, Smyth P, Finn SP, Denning K, Flavin R, O'Regan EM, Li J, Potratz A, Guenther SM, Henfrey R, O'Leary JJ, Sheils O. Effect of ret/PTC 1 rearrangement on transcription and post-transcriptional regulation in a papillary thyroid carcinoma model. Mol Cancer. 2006; 5:70. https://doi.org/10.1186/1476-4598-5-70 [PubMed]
- 28. Bonetti P, Climent M, Panebianco F, Tordonato C, Santoro A, Marzi MJ, Pelicci PG, Ventura A, Nicassio F. Dual role for miR-34a in the control of early progenitor proliferation and commitment in the mammary gland and in breast cancer. Oncogene. 2019; 38:360–74. https://doi.org/10.1038/s41388-018-0445-3 [PubMed]
- 29. Hanieh H, Ahmed EA, Vishnubalaji R, Alajez NM. SOX4: Epigenetic regulation and role in tumorigenesis. Semin Cancer Biol. 2020; 67:91–104. https://doi.org/10.1016/j.semcancer.2019.06.022 [PubMed]
- 30. Lu JW, Hsieh MS, Hou HA, Chen CY, Tien HF, Lin LI. Overexpression of SOX4 correlates with poor prognosis of acute myeloid leukemia and is leukemogenic in zebrafish. Blood Cancer J. 2017; 7:e593. https://doi.org/10.1038/bcj.2017.74 [PubMed]
- 31. Paul MH, Harvey RP, Wegner M, Sock E. Cardiac outflow tract development relies on the complex function of Sox4 and Sox11 in multiple cell types. Cell Mol Life Sci. 2014; 71:2931–45. https://doi.org/10.1007/s00018-013-1523-x [PubMed]
- 32. Malhotra N, Qi Y, Spidale NA, Frascoli M, Miu B, Cho O, Sylvia K, Kang J. SOX4 controls invariant NKT cell differentiation by tuning TCR signaling. J Exp Med. 2018; 215:2887–900. https://doi.org/10.1084/jem.20172021 [PubMed]
- 33. Ding L, Zhao Y, Dang S, Wang Y, Li X, Yu X, Li Z, Wei J, Liu M, Li G. Circular RNA circ-DONSON facilitates gastric cancer growth and invasion via NURF complex dependent activation of transcription factor SOX4. Mol Cancer. 2019; 18:45. https://doi.org/10.1186/s12943-019-1006-2 [PubMed]
- 34. Wang K, Long B, Zhou LY, Liu F, Zhou QY, Liu CY, Fan YY, Li PF. CARL lncRNA inhibits anoxia-induced mitochondrial fission and apoptosis in cardiomyocytes by impairing miR-539-dependent PHB2 downregulation. Nat Commun. 2014; 5:3596. https://doi.org/10.1038/ncomms4596 [PubMed]
- 35. Paraskevopoulou MD, Vlachos IS, Karagkouni D, Georgakilas G, Kanellos I, Vergoulis T, Zagganas K, Tsanakas P, Floros E, Dalamagas T, Hatzigeorgiou AG. DIANA-LncBase v2: indexing microRNA targets on non-coding transcripts. Nucleic Acids Res. 2016; 44:D231–38. https://doi.org/10.1093/nar/gkv1270 [PubMed]
- 36. Zhao C, Mo L, Li C, Han S, Zhao W, Liu L. FOXN3 suppresses the growth and invasion of papillary thyroid cancer through the inactivation of Wnt/β-catenin pathway. Mol Cell Endocrinol. 2020; 515:110925. https://doi.org/10.1016/j.mce.2020.110925 [PubMed]