




Introduction
Low back pain (LBP) is one of the most common musculoskeletal conditions requiring medical care and contributing to patient impairment, disability, and enormous costs [1, 2]. A primary cause of LBP is intervertebral disc degeneration (IDD), characterized by progressive depletion of matrix proteoglycan resulting in disc height reduction, altered biomechanics, and possible nerve compression and pain [3–5]. Age is by far the leading risk factor for IDD [6, 7]. Aged intervertebral discs (IVD) and concomitant loss of tissue function are often associated with the accumulation of senescent cells (SnCs) [8, 9]. Senescence is a cell fate characterized by stable growth arrest in response to stress and damage and acquisition of an inflammatory senescence-associated secretory phenotype (SASP). The SASP is characterized by chronic secretion of a variety of inflammatory cytokines, chemokines and proteases that can disrupt tissue structure and function. A key role for cellular senescence in driving aging and age-related diseases has been demonstrated using both genetic and pharmacological approaches to reduce the senescent cell burden. For example, clearance of SnCs improves the biological function of cells and tissues, including the brain [10], heart [11], skin [12, 13], skeletal muscles [14], cartilage [15] and bone [16], and extends the healthspan of mouse models of aging [12, 17, 18].
The number of SnCs increases with aging and degeneration in the IVD of humans and animal models [5, 19, 20]. This increase is especially pronounced in post-menopausal females or ovariectomy in rodent models [21]. In aged IVD, matrix proteoglycan loss is highly associated with an increased SnC burden [5], which is attributed to the action of the SASP [9, 22]. IVD SnCs produce elevated levels of matrix metalloproteinases such as MMP-13 and ADAMTS-4 and the pro-inflammatory cytokines IL1β, TNFα, IL6 and IL8, all of which are thought to be a part of the SASP of senescent disc cells [5]. These SASP factors are expressed at higher levels in post-menopausal women, possibly explaining the higher rate of self-reported chronic low back pain and clinically evident IDD in older women compared to men [23, 24]. Together, these SASP factors are postulated to promote degradation of the existing IVD matrix and prevent regeneration through the propagation of cellular senescence and the SASP to neighboring cells via paracrine or endocrine mechanisms [5, 25]. A defining characteristic of SnCs is their resistance to apoptotic cell death [26, 27]. SnCs escape apoptosis by upregulating various “senescent cell anti-apoptotic pathways” (SCAPs), including the p38 MAPK, mTOR and BCL-2 family anti-apoptotic pathways [18, 27]. Among these, overexpression of the BCL-2 family anti-apoptotic proteins BCL-xL/BCL-2 is strongly linked to SnCs survival, and blocking this anti-apoptotic target results in selective elimination of SnCs.
Senolytics are compounds that selectively remove SnCs by counter-acting SCAPs. Removal of SnCs with senolytics rejuvenates tissues in vivo and prolongs healthspan in animals [18, 28]. The senolytic drug combination of Dasatinib and Quercetin (D+Q), which inhibit src/tyrosine kinases and the anti-apoptotic BCL-2 and other targets, respectively, has also been shown to reduce IDD [25, 29]. Recently, the BCL-2/BCL-xL dual inhibitor ABT-263 (Navitoclax) was shown to have senolytic activity and reduce disc degeneration after intervertebral disc puncture using a novel polylactic-co-glycolic acid (PLGA) nanoparticle delivery system during the injury [30]. However, ABT-263 also suffered from platelet toxicity, reducing its utility in fighting cancer, for example. To overcome enhance the activity of ABT-263 and reduce its toxicity, the von Hippel-Lindau (VHL) E3 ligase was conjugated to ABT-263 to create PROTAC 753b (753b), targeting the complex for ubiquitination and proteolysis [31]. Because platelets have relatively low VHL, the toxicity in these cells observed for ABT-263 was expected to be limited.[32] Indeed, the PROTAC 753b showed greater efficacy in the destruction of senescent cells while demonstrating decreased platelet toxicity during treatment of leukemia [33], small cell carcinoma [34], and hepatocellular carcinoma [31, 32]. The properties of 753b suggest that it can function as a more potent senolytic with less platelet toxicity than ABT-263 and other BCL-2/BCL-xL inhibitors. Indeed, we recently reported that 753b can effectively clear SnCs in the liver and reduces metabolic dysfunction-associated steatohepatitis-driven hepatocellular carcinoma in mice [35].
Despite its superior senolytic properties as compared to ABT-263, 753b has not yet been tested for its ability to alleviate age-related IDD. In this study, we assessed whether systemic 753b treatment can reduce age-related IDD in mice. Here we treated 16-month mice with 753b over a 6-month period by intraperitoneal (IP) injection 2x/week for 2 weeks per cycle followed by a two-week period of no treatment. Although blood vessels do not penetrate the nucleus pulposus or inner annulus fibrosus, capillary networks terminating within the vertebral bodies and subchondral bone immediately adjacent to the cartilaginous endplates provide a pathway for small molecule diffusion into the nucleus pulposus [36]. Systemic delivery of 753b was anticipated to allow entry of 753b into the NP of the IVD and change its activity based on the diffusion properties of glucose, oxygen, lactate, contrast agents and PET markers (i.e., 18F-FAPI-04) as well as antibiotics like cephazolin and nutraceuticals, which are substantially larger than PROTAC 753b. The concentration of these molecules has been shown to reach disc:plasma ratios of 25-50% within 6 hours of systemic delivery [37–42]. In this study, the IVDs of 753b- and vehicle-treated old mice and 6-month untreated young control mice (n=5 in each group) were assessed by histology, immunohistochemistry, Western blot and gene expression of key IVD senescence markers.
Although this study was originally designed to evaluate systemic senescence modulation, spine tissues were analyzed after study completion as a biologically relevant, age-sensitive organ system. In this study, we hypothesized that 753b would reduce age-related IDD. The IVDs of male and female 753b- and vehicle-treated old mice as well as 6-month untreated young control mice (n=5 in each group) were assessed by the MERCY histomorphometric scoring system, immunohistochemistry for the targets of 753b within the IVD tissue and inflammatory markers in the blood, Western blot for BCL2 and BLC-XL and matrix degradation products in the IVD, and gene expression of key senescence markers. We reasoned that this collection of data would identify positive effects of aged IVD tissues to the senolytic 753b, serving as hypothesis generating if not indicative of the effectiveness of 753b and BCL2 family targeting PROTACS in reducing age-related IDD.
Results
753b treatment improved intervertebral disc histological features in aged male mice
Given the reported causative role of cellular senescence in IVD aging pathology and disease progression, we treated 16-month C57BL/6J mice with a regimen of 753b illustrated in Figure 1C to remove SnCs pharmacologically, as we recently reported [35]. This treatment involved IP injection of 753b 2x/week for 2 weeks per cycle with an interval of two weeks of rest between the cycles for the duration of the experiment (Figure 1C). Using the MERCY histological scoring system for mouse IDD [43], we observed greater degeneration in all disc tissue compartments in both 22m old male and female mice treated with vehicle as compared to young 6m old mice (Figure 2). However, old male mice had more significant IDD pathology with advanced age than females, with females generally exhibiting significantly less markers of degeneration at baseline in all IVD tissues except the NP. Among males, we observed a total increase in MERCY score of 10.67 ± 1.89, p < 1.27 x 10-7) as compared to 4.56 ± 1.77, p < 8 x 10-3 from 6-month to 22-month-old mice. In males, there was an average increase of 0.83 + 0.24 in degeneration scale across all tissues of the IVD including the nucleus pulposus (NP), annulus fibrosus (AF) and endplate (EP), whereas females exhibited a more modest level of degeneration (0.41 + 0.28) when comparing 22m old mice with 6m old mice, possibly as a function of their healthier baseline.
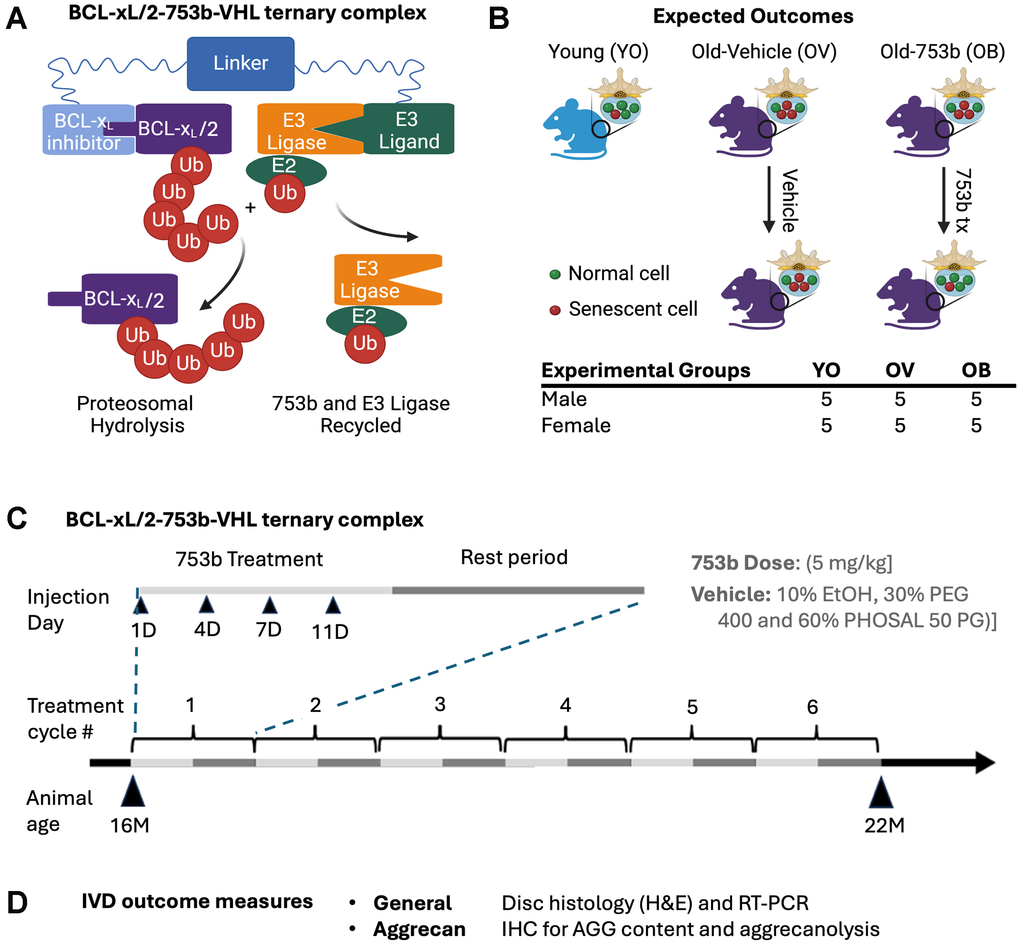
Figure 1. Schematic of study design. (A) PROTAC 753b consists of a BCL-xL/2 binding moiety connected to a Von Hippel-Lindau (VHL) E3 ligase ligand via a linker. 753b functions by binding to BCL-XL/BCL-2, targeting them both to the VHL E3 ligase for ubiquitination and subsequent proteasomal degradation. Treatment of mice (B) with 753b via IP injection through a treatment regimen (C) consisted of 6 1-month treatment cycles comprised of a 2-week drug delivery period followed by a 2-week rest period. During the treatment period, the drug was delivered 2 times per week. Peritoneal injections of 753b (5 mg/kg) or vehicle control (10% ethanol, 30% PEG 400 and 60% PHOSAL 50 PG) began at 16 months of age (16M) ending at 22 months of age (22 M) for a total of 6 months. Vehicle (10% ethanol, 30% PEG 400 and 60% PHOSAL 50 PG), 753b (5mg/kg 753b). (D) Outcome measures of harvested IVDs from the spines.
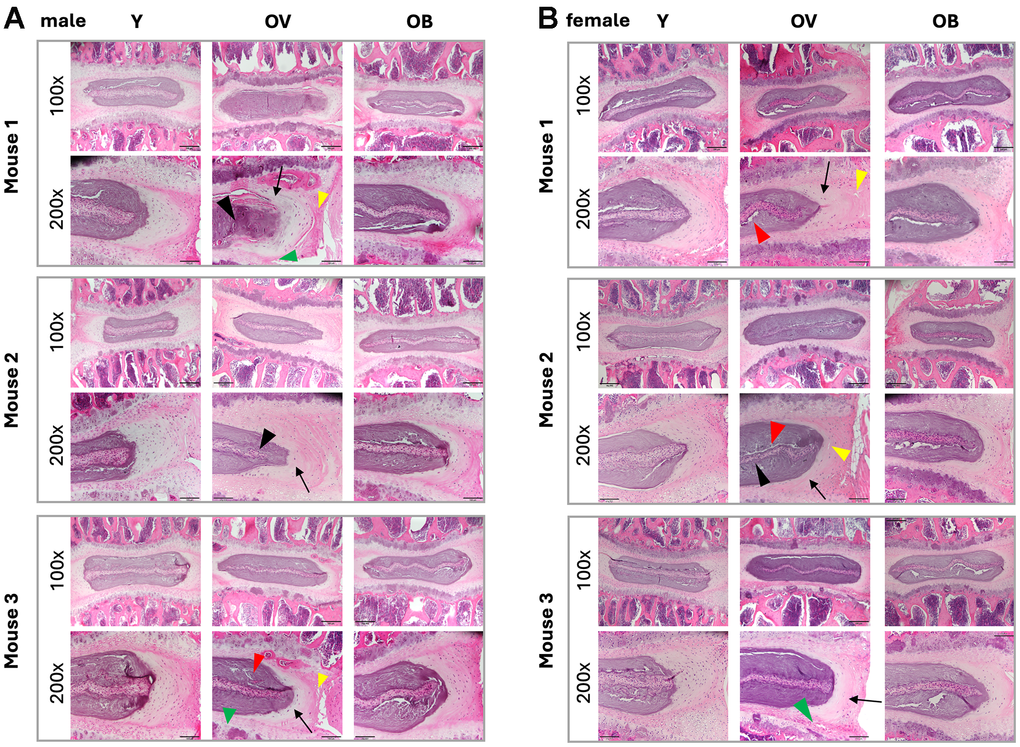
Figure 2. Effect of aging and 753b treatment on mouse IVD histomorphology. Serial sections of three representative (A) male and (B) female mice stained with H&E and imaged at 100x and 200x magnification for MERCY scoring. Labels: Y, young 6m animals, OV, 22m animals in the vehicle treatment group, OB, 22m animals in the 753b treatment group. Note the decrease in cellularity, matrix organization, and tissue integrity (e.g., increase in fissuring) in the different tissues of the aged 22m animals versus the young 6m animals. Legend: Black arrows, NP/AF boundary; yellow arrowheads, loss of AF structure (serpentine lamellae widely spaced); red arrowheads, loss of NP matrix indicated by large empty space gaps; black arrowheads, NP fissures and clefts= Black arrowheads; green arrowheads, presence of severe microfracture and Schmorl’s node in the EP.
MERCY scoring of the 753b treated mice revealed a significant positive benefit on IVD in aged males, but not females (Figure 3). Among males, there was a total reduction in the MERCY score of 5.05 ± 3.83, p < 0.022 with an average decrease of 0.45 ± 0.14 in degeneration scale across all tissues following 753b treatment (Supplementary Table 1, Supplementary Materials). However, among the male aged cohorts, no significant improvements (i.e., decreases in MERCY score) with 753b treatment were seen in any individual IVD tissue compartment. In aged females, 753b treatment improved NP health, but none of the other disc compartments, including AF, EP, and AF/NP/EP interfaces, resulting in a composite MERCY score that is not statistically significant in female mice compared to vehicle control (Figure 3).
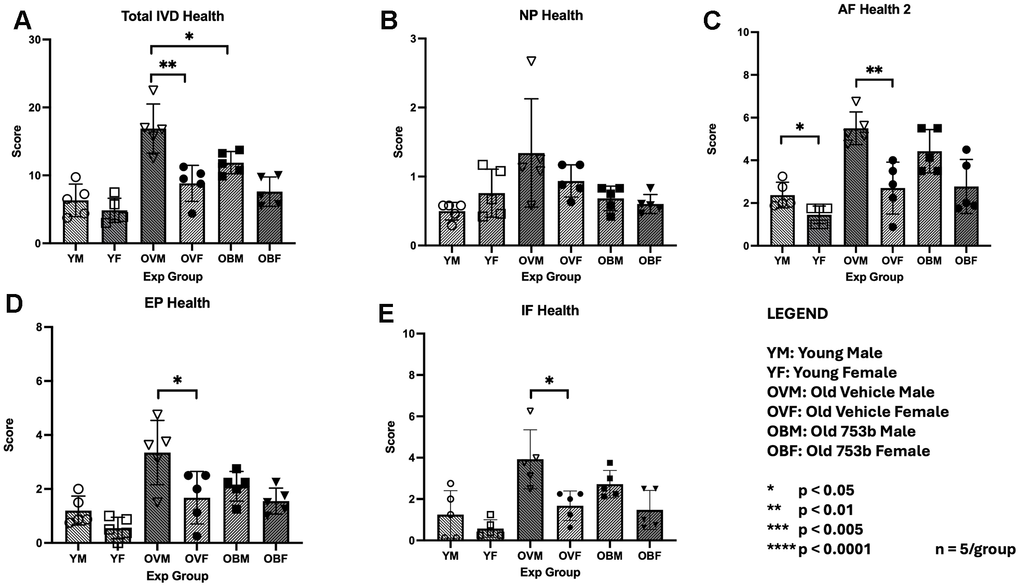
Figure 3. Histomorphological changes in the IVD tissues with age and 753b treatment. MERCY scores for (A) all tissues of the IVD together (composite) and individual tissues of the IVD that comprise the MERCY score: (B) nucleus pulposus (NP), (C) annulus fibrosus (AF), (D) endplate (EP), and (E) AF/EP interface. Scores were expressed as mean+SEM for n=5 per mouse group. * Denotes statistical significance in response to 753b treatment.
753b treatment improved NP aggrecan content in IVDs of aged male mice
Aggrecan is the major extracellular proteoglycan matrix molecule responsible for maintaining the hydrostatic pressure within the disc nucleus pulposus. Aggrecan loss and degeneration of the nucleus pulposus are often the first and most conspicuous changes in IDD. In male mice, semi-quantitative immunohistochemistry for aggrecan revealed a decrease in NP aggrecan (54 ± 5%, p < 0.00183) in the aged 22m mice versus young 6m controls (set at 100%) (Figure 4A). 753b treatment increased NP aggrecan to 50% ± 23% (p < 0.0384) compared to the old vehicle-treated males (Figure 4A, 4B), returning the NP aggrecan staining to a level similar to that of the young males. In the female cohort, age-dependent NP aggrecan loss was not observed in 22m old mice compared to 6m young mice, and 753b treatment had no significant effect on NP proteoglycan content (Figure 4C, 4D).
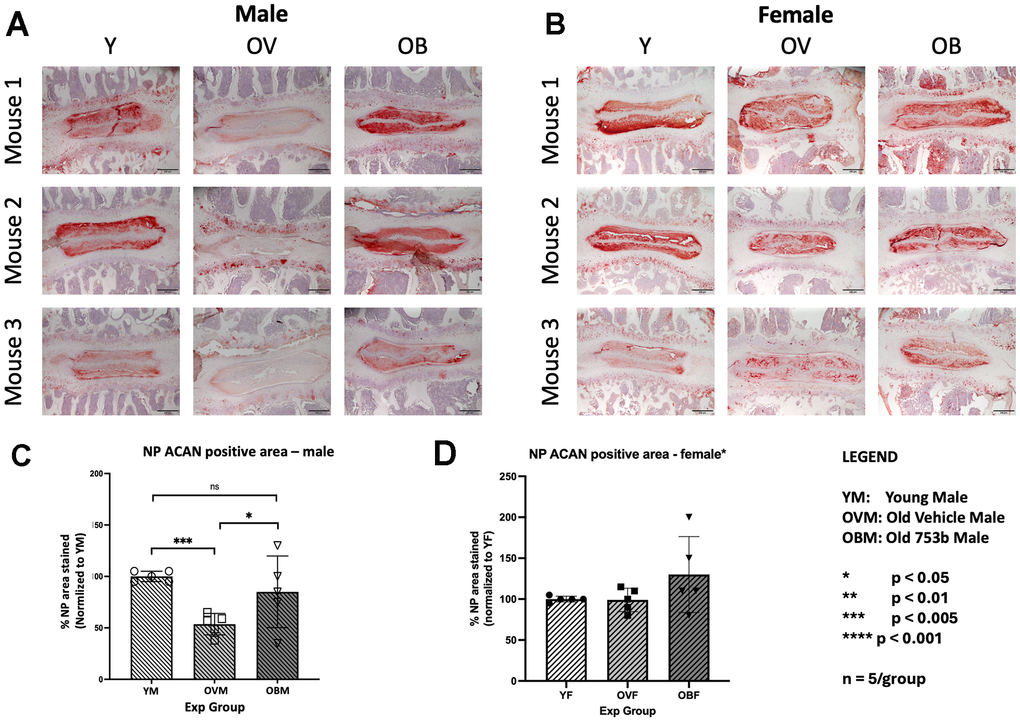
Figure 4. Effect of aging and 753b treatment on mouse IVD tissue aggrecan protein production. Serial sections of three representative (A) male and (B) female mice processed to visualize aggrecan production imaged at 100x. Quantitation of aggrecan staining by Image J particle analysis in (C) male and (D) female mice showing average values of n=5 mice/group ± SEM.
Effect of 753b treatment on IVD tissue aggrecan degradation
Loss of aggrecan can be caused by reduced anabolism, increased catabolism or both. Aggrecan degradation due to cleavage within its interglobular domain (IGD) between G1 and G2 is considered most pathological because it leads to loss of the entire GAG-containing region of aggrecan that is vital for disc function [44]. To assess disc aggrecan breakdown, Western blots were performed to detect MMP- and ADAMTS-mediated proteolytic cleavage within the IGD of aggrecan (Figure 5A, 5B). Quantitation of the Western blots revealed large increases in MMP-mediated cleavage products (Figure 5C, 5D) in both aged males (≈ 4-fold increase, p<0.0043) and females (≈ 3-fold increase in female mice, p<0.0069) as well as ADAMTS-mediated cleavage products, which increased ≈ 4-fold in aged males (p < 0.0002) and ≈ 2.5-fold in aged females (p < 0.0002). With 753b treatment in male mice, the MMP-generated aggrecan fragments quantity trended lower (p < 0.055) but did not attain significance. In the female cohort, 753b had no significant effect on reducing the levels of these IVD aggrecan fragments in aged mice.
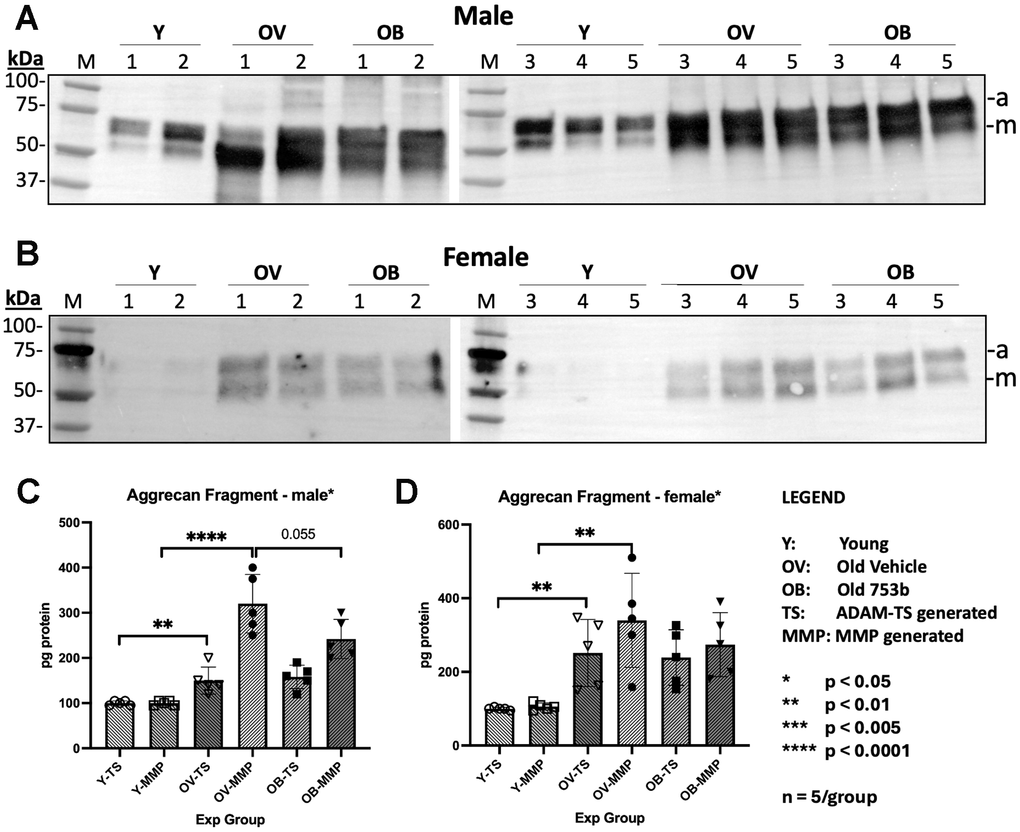
Figure 5. Effect of aging and 753b treatment on aggrecan protein fragmentation in mouse IVD tissues. Western blots for aggrecan in total protein of whole IVDs isolated from five (A) male and (B) female animals. Quantitation of the Western blot results of (C) male and (D) female for aggrecan and MMP-generated fragments in total protein of IVDs by the Bio-Rad GelDoc. a, ADAMTS-generated aggrecan fragments; m, MMP-generated aggrecan fragments.
Effect of 753b treatment on IVD cellular senescence
753b induces apoptosis in SnCs by degrading the anti-apoptotic proteins BCL-2 and BCL-xL [35]. To assess the senolytic activity of 753b treatment in IVD, we used RT-PCR to assess the expression of key biomarkers of cellular senescence. Our data showed gene expression of cell cycle regulator p16INK4 was elevated with age in the IVD of female rats (p < 0.0004) but not in males (Figure 6A). Surprisingly, 753b treatment failed to reduce the increased levels of p16INK4 mRNA level in both sexes. In fact, 753b treatment significantly increased p16 over 6-fold in males (p < 0.0214) and females (p < 0.0346). mRNA expression of several key SASP factors, IL-6, IL-8, and TNFα generally trended toward increase with age in both sexes, but significance in that increase was only observed among females for IL8 (p < 0.030) and TNFα (p < 0.021). Female rats also had significantly lower baseline expression of IL8 (p < 0.009) than male rats (Figure 6B–6D). Together, these results suggest little or no effect of 753b on cellular senescence burden in the IVDs of the 753b-treated mice. This finding agrees with the results from our previously reported study [35] that showed differing reductions of SnCs in the liver, lung, kidney and fat tissues in aged mice in part due to its differential tissue distribution. By clearing SnCs in the liver and other tissues, 753b reduced IL-6 and TNFα protein levels in the plasma of treated male mice, suggesting that the benefits on male discs by this senolytic might be attributable to systemic effects rather than the local ones. This suggestion is supported by our previous studies in which we found that systemic treatment with ABT-263, which does not cross the blood-brain-barrier, still alleviated doxorubicin-induced senescence via reducing plasma SASP factors in rats [35].
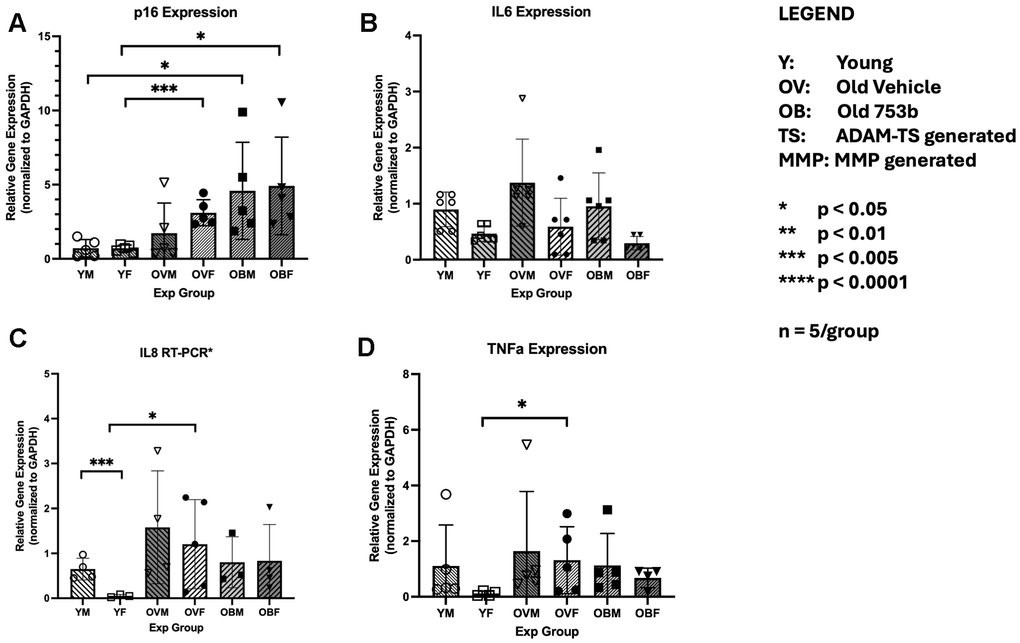
Figure 6. The effect of aging and 753b treatment on gene expression of key markers of cellular senescence in mouse disc tissue. Shown are the RT-PCR results for the cell cycle regulator Cdkn2a for p16INK4 (A), and the SASP factors IL-6 (B), IL-8 (C) and TNFa (D). n=5/group (each run in triplicate).
Discussion
Cellular senescence in IVD aging and degeneration
We and others have demonstrated that the percentage of SnCs in the IVD is increased during IVD aging and degeneration in both human and animal models [25, 45, 46]. We previously demonstrated a strong association between cellular senescence and disc proteoglycan (PG) matrix loss in various mouse models of age-dependent IDD, including natural aging in wild-type mice, accelerated aging in the Ercc1-/∆ mouse model of a human progeroid syndrome due to DNA repair-deficiency, and mice chronically exposed to genotoxic agents such as ionizing radiation, mechlorethamine, or tobacco smoke. Likewise, senescent disc cells in culture exposed to genotoxic agents such as cisplatin, H2O2, or ionizing radiation acquire SASP, lose their PG matrix homeostasis, and exhibit enhanced proteoglycan catabolism and reduced anabolism [47–49]. Genetic and pharmacologic reduction of signaling activity of NF-κB, a central mediator of SASP establishment and inflammation, restored disc PG homeostasis in the Ercc1-/∆ mouse model of accelerated aging. Moreover, depletion of p16-positive SnCs in the transgenic p16 INK4a-3MR mice mitigated disc PG loss. Together, these studies provide compelling evidence supporting the causative roles of persistent DNA damage and cellular senescence in driving age-related IDD, suggesting pharmacologic approaches to clear SnCs would be therapeutic, which we and others have observed with treatment with the senolytic combination of D+Q [25, 50].
D+Q has consistently been shown to reduce the senescence burden in aged IVD tissues concomitant with lower levels of degeneration, significant decreases in senescence markers p16 and p19, and reductions in SASP molecules IL-6 and MMP13 that contribute to improved cell viability, matrix content and tissue morphology [25]. The same group subsequently compared D+Q treatment with ABT-263 (Navitoclax) treatment and found that ABT-263 did not improve disc degeneration in SM/J mice [51], suggesting that the efficacy of ABT-263 may be limited to techniques of local administration or possibly dependent on the local concentration of the drug as previously employed in the prevention of disc degeneration [30]. The novel design of PROTAC 753b, with its improved efficacy and lower toxicity, allows application of higher doses, promising a systemic application of this senolytic to prevent age-related disc degeneration. Despite observing more subtle effects on age-related disc degeneration, we were unable to detect any change in BCL-2/BCL-xL in the 753b treated discs. However, reductions in serum inflammatory markers indicate that systemic inflammation may be reduced in the 753b treated mice and that this positively impacts IVD health though indirect means. In contrast, Risbud et al. were able to show specific targets of D+Q treatment, ephrin B (Dasatinib) and the PI3K/AKT (Quercetin) pathways within the IVD [51]. The context or conditions under which 753b treatment may be effective, i.e. well-perfused, metabolically active (endo/exocytic) tissues that exist in lung, liver and tumors, do not exist in the IVD. This emphasizes the idea that the efficacy of senolytics is highly context-dependent.
753b effect on IVD health
In this study, we tested the effect of systemic delivery of the BCL-2/BCL-xL targeting PROTAC 753b, which does not have the same platelet toxicity caused by the first-generation dual BCL-2/BCL-xL inhibitor navitoclax (ABT-263) [31, 33]. Interestingly, our results demonstrate that the PROTAC 753b can significantly slow age-associated IDD in male but not female mice. In addition, 753b clearly improved the histomorphometry of the IVD in aged male mice with improvements in all IVD tissue compartments, including AF, EP, NP/AF/EP interface, and to a lesser extent NP. IVD aggrecan homeostasis, assessed by measuring total aggrecan protein content and aggrecanolysis, was better maintained in 753b-treated than vehicle-treated male mice, consistent with the improvement in IVD histomorphology. Compared to vehicle, IVDs of 753b-treated aging male mice exhibited less age-related aggrecan loss and MMP-mediated aggrecan proteolysis. Senescent disc cells upregulate both MMPs and ADAMTSs as part of their SASP [46, 52]. It is possible the relative induction of these enzyme families, and the resulting aggrecan fragment production, is a result of differential abundance of these enzymes in the IVD or that microenvironmental or sex-specific factors contribute to the distinct patterns of catabolic enzyme function. Direct evidence for preferential MMP versus ADAMTS expression in senescent disc cells is currently lacking and needs to be investigated. The consequences of the observed differential activity are interesting. Aggrecanase (ADAMTS) activity is closely linked to aggrecan/proteoglycan catabolism, which reduces matrix osmotic swelling pressure, water content [53, 54], and thereby nucleus pulposus hydrostatic pressurization, whereas MMPs are widely recognized as major regulators of ECM remodeling (including collagenous matrix turnover [55]. 753b may be acting to stabilize the ECM through collagen network preservation, but not preserving the Aggrecan network and thus not improving the hydrostatic function of the tissue, rendering the disc still vulnerable to degeneration.
We have previously observed similar beneficial effects of senolytics in natural aging mice and progeroid Ercc1-/∆ mice treated with a combination of D+Q [25, 50], although these studies did not distinguish if D+Q confer benefits to IVD health equally or differently between female and male mice. Likewise, systemic delivery of several other senolytics has been reported to ameliorate IDD following AF puncture in mouse models. These senolytics include Quercetin, ABT-263, and RG-7112 [25, 56–58] which were reported to preserve IVD structure through expression and retention of key matrix molecules such as aggrecan and collagens. In IVD cell culture models, the natural compound curcumin and its metabolite, o-vanillin were postulated to have senolytic action as they reduced the number of p16-positive cells and increased the number of Ki-67 and caspase-3-positive cells in monolayer and pellet cultures [59].
The sexual dimorphism of 753b effects
It is intriguing that in this study 753b maintains IVD health in aging male mice, but not female mice. Such sexual dimorphism has been observed for other age-associated therapeutic treatments. Lifespan extension drugs such as 17 alpha estradiol (17aE2), nordihydroguaiaretic acid (NDGA), and aspirin have been shown to extend longevity only in males [60, 61]. Though rapamycin and acarbose increased longevity in both sexes, the effect of acarbose was much greater in males [60], while the effect of rapamycin was greater in females [62]. Estrogen may account for some of these results, and studies in mice have demonstrated that male lifespan can be extended with an estrogenic agonist [63]. In our study, it is unclear why the therapeutic effect of 753b is more apparent in IVDs of aging male than female mice. Here, aging male mice showed a much greater degree of age-dependent IDD than female mice at the age of 22 months when compared to young mice of 6 months old. It is conceivable that our 753b treatment reduced IDD in aged males but not aged females in part because female mice at 22 months of age have yet developed noticeable age-related IDD, possibly due to a lower level of senescence in the IVD. This is in contrast to the human condition, in which post-menopausal females suffer higher rates of IDD compared to male counterparts that are coincident with higher SASP levels and increased senescence in the IVD. Female mice are known to lack a true menopause, and this fact alone may account for the observed inability of 753b to significantly reduce IDD in aged mice. Further studies are needed to understand the sexual dimorphism effects of 753b on IVDs.
Mechanism of action of 753b on improving IVD health
It is interesting that we observed clear histomorphogenic changes in aged male rats treated with 753b in the absence of any changes in the immediate targets of 753b, BCL2 or BCL-xL. Because the mechanism of 753b is known to impact senescence directly, the observed therapeutic effects of 753b on IVDs of the male mice in the absence of changes in IVD senescence markers suggest a global reduction of cellular senescence burden through systemic, non-cell autonomous processes. Previous research from our lab and others demonstrate that cellular senescence has a causal role in driving disc aging, but whether it does so locally through cell autonomous, systemically via non-cell autonomous mechanisms, or both is still poorly understood. Specifically, it is not known if cells within the IVDs become senescent and accumulate over time in aging animals, which then cause disc PG homeostatic imbalance and age-associated IDD in a local cell autonomous manner, or systemic SASP factors secreted by SnCs in other tissues of aging animals contribute to inflammation and PG imbalance in disc tissue via a non-cell autonomous pathway. These two pathways cannot be distinguished in the animal models previously used to study disc aging because DNA damage and cellular senescence were elicited systemically in those models. In addition, our studies of elimination of SnCs in p16 INK4a-3MR transgenic mice by a genetic strategy and in Ercc1-/∆ mice and natural aging mice by senolytic treatments were performed exclusively by systemic clearance, making it impossible to distinguish if mitigation of age-dependent IDD observed in those studies was due to solely to local clearing of senescent disc cells or in combination with systemic clearance of cellular senescence.
The liver and spleen accumulated much higher levels of 753b for a long duration than the lung, kidney, and fat tissues [35]. It is possible that the senolytic 753b enhances IVD health by directly clearing the SnCs in aged mouse IVD tissue, thereby eliminating a presumably key driver of age-related IDD. If this is true, 753b from the systemic treatment via IP injection is expected to penetrate the IVDs and promote degradation of the anti-apoptotic protein BCL-2/BCL-xL in order to clear the resident IVD SnCs. Indeed, IVD cellular senescence burden is elevated in our aged mice, evidenced by increased expression of key senescence markers IL-6, IL-8, TNFα and p16INK4a in 22m old mice compared to 6m old mice (Figure 6) [64]. However, 753b treatment did not reduce expression of these senescence markers in IVD tissue. Instead, plasma protein levels of IL-6 and TNFα were decreased in old male mice treated with 753b (Supplementary Figure 1), suggesting that the therapeutic action of 753b on aged mousse IVD is in part systemic and not through local removal of senescence cells from the IVD. Consistent with this result is the fact that we did not see changes in BCL2 abundance, the anti-apoptotic protein target of 753b, in IVDs of 22-month-old mice (Supplementary Figure 2). These results strongly indicate that 753b blunts age-related IDD indirectly through targeting SnCs systemically. This suggestion is supported by our previous findings that 753b treatment can effectively reduce senescent cell burden in the liver and spleen in old mice [35].
Recent evidence suggests disc aging and degeneration may be influenced by factors beyond the local disc environment. For example, osteopenia and osteoporosis correlate with increased incidence of IDD in which sclerosis of the endplate reduces the nutrient transport to disc tissue [21, 65]. In addition, muscular dystrophy has been shown to reduce PGs in the IVD leading to acceleration of degenerative disc disease in a Duchenne muscular dystrophy model [66]. Our heterochronic parabiosis pairing of young and old mice showed that exposing young mice to old blood accelerated their disc matrix imbalance and tissue degeneration, demonstrating non-cell autonomous effects of circulating factors on disc aging [67]. Moreover, an aged senescent immune system has a causal role in driving systemic ageing, including age-related IDD. In addition, we previously reported that systemic treatment with ABT-263, which does not cross the blood-brain-barrier, could alleviate doxorubicin-induced chemobrain by reducing plasma SASP factors in rats [35]. Thus, it appears that disc aging is often driven, in part, by non-cell autonomous pathways and global cellular senescence, which may be reduced by 753b systemic treatment to result in improved IVD health in our 22m old male mice.
Study limitations
This study was originally designed to evaluate the systemic effects of a senescence-targeting therapeutic on age-associated pathology across multiple organ systems. Intervertebral disc tissue was analyzed as a secondary endpoint to explore whether systemic modulation of aging biology is reflected in an avascular, slowly remodeling musculoskeletal tissue. This culminates in several limitations. First, although the pharmacokinetics had been determined for several soft tissues [35, 66, 68], similar studies for skeletal tissues such as the spine were not conducted. Thus, it is unclear whether 753 reached in the IVD and entered the cells to modify apoptosis of SnCs directly. The dosing in this study was determined based upon 753b bioavailability to other vascularized, soft tissues and aligned with diffusion of many molecules occurring in under 6 hours with a disc-to-plasma ratio of 25-50%. However, this diffusion is highly dependent on the solute properties [69]. In antibiotics research, high net positive charge prevents bioavailability to NP and reduces NP membrane permeability of the solute. The net change of PROTAC 753b at pH 6.8-7.4 is estimated to be +5. At this level, membrane permeability is greatly reduced and partitioning to the extracellular matrix is greatly increased. The disc is maybe least ideal for PROTAC delivery (and subsequent function) because it is avascular and diffusion-limited, highly anionic (proteoglycan-rich), metabolically quiescent and exists at a relatively low pH. In addition, permeability, diffusion and metabolic activity worsen with age.
A second limitation stems from the small group size, which had consequences in several key parts of this study, especially in assessing the mechanism of 753b action in aged male rats. The low availability of samples resulted in limited opportunities to examine specific targets of 753b. Immunohistochemistry for BCL2 and BCL-xL revealed low signal in the IVD tissues and no difference between 753b-treated and control individuals. We chose not to assay apoptosis directly (by TUNEL) or caspase 3 cleavage instead because changes in the rate of apoptosis between vehicle and 753b treated animals would be hard to detect given the cell number in the NP is low and the fraction undergoing apoptosis significantly lower. A full characterization of senolytic pathways at the protein level was also not possible. Where trends in senolytic gene expression or plasma protein were noted, a larger group size may have resulted in a more definitive conclusion and put a finer point on the observed sexual dimorphism. Together, these limit the findings reported as more hypothesis-generating rather than mechanistic.
Despite these shortcomings, the results of this study, including improvement in IVD histomorphometry and MMP-mediated matrix remodeling in aged male rats, suggest that 753b does ameliorate age-related IDD in male rats. These findings suggest that the BCL-2/BCL-xL targeting PROTAC 753b can ameliorate age-related IDD, representing a viable therapy strategy for mitigating IDD associated with aging.
Materials and Methods
Mice, treatment, and study design
In the execution of this study originally designed to evaluate systemic senescence modulation, spine tissues were analyzed after study completion as a biologically relevant, age-sensitive organ system. C57BL/6J mice were randomized and treated intraperitoneally with 753b (5mg/kg) or Vehicle (10% ethanol, 30% PEG 400 and 60% PHOSAL 50 PG), 5 mice per group. This experimental group size was deemed likely consistent based on previous work by others published at the time of study design indicated that an n of 3-5 may be sufficient to record differences [65, 66, 70, 71] to distinguish positive effects of aged IVD tissues to senolytics and the results could at least serve as hypothesis generating if not indicative of the effectiveness of 753b and PROTACS in reducing age-related IDD. The 753b treatment occurred over a 6-month period starting at 16 months of age and ending at 22 months of age, when the mice were sacrificed for analysis (Figure 1). 16-month-old and 22-month-old mice are equivalent to middle-aged (47 year) and old (60-70 year) humans [72, 73]. 6-month mice were used as a comparative control representing a young adult with healthy intervertebral discs to help address age-related directionality of changes [74, 75]. The dose of 753b was selected based on a preliminary pharmacokinetics study, as done for treatment of murine puncture-induced IDD with ABT-263 reported previously [76, 77], designed to identify a dose of 753b highly potent for BCL-XL and BCL-2 degradation [33]. 753b or vehicle alone was administrated twice a week (Monday and Thursday) for 2 weeks per cycle with a two-week inter-cycle resting period (Figure 1C). Mouse health and weight were recorded weekly. Five animals in each of the following six cohorts were treated and sacrificed for IVD assessment: aged male vehicle control, aged male 753b-treated, young male comparative, aged female vehicle control, aged male 753b-treated, and young female comparative. RT-PCR and western blots were performed on the 10 most proximal coccygeal (tail) discs. We used lumbar discs for morphologic outcome validity (histologic degeneration scoring in the clinically relevant region) and coccygeal discs for mechanistic sensitivity, where RNA/protein endpoints require higher tissue yield and tighter time-course control. In doing this, we acknowledge that lumbar and tail discs might have some structural and mechanical differences. But in dividing the analysis types by disc level, we are matching assay-to-tissue outcomes while not claiming the discs are the identical.
Histological analysis
L3–L5 lumbar segments isolated from aged (22m) male and female 753b-treated and vehicle controls and that of untreated healthy, young (6m) males and females were fixed in 10% formalin and decalcified in Decalcifier I solution (3800440, Leica) at 4° C overnight. All tissues were dehydrated through a graded alcohol series and then embedded in paraffin and cut into 4-μm thick sections in the coronal plane. Serial sections were stained with hematoxylin and eosin (H&E; HT110280-2.5L, HHS128-4L, MilliporeSigma) by standard procedures and then photographed under 100 or 200× magnification (E800 microscope, Nikon Eclipse) [45, 78]. Degenerative changes in the mouse IVD of aged mice were assessed using the MERCY histological scoring system [43] for 5 animals in each cohort (Figure 1B). Four scorers blinded to the mouse IDs scored the H&E stained sections from each mouse for 14 specific features in the nucleus pulposus (NP), annulus fibrosus (AF), cartilaginous endplate (EP), and NP/AF interface to calculate a composite score representing the total of all these 14 individual scores (Supplementary Table 1, Supplementary Materials). The features of NP and AF that were scored included NP cellularity and morphology, NP fibrosis, NP matrix organization, AF cellularity, AF bulging, AF lamellar organization, and AF clefts/fissures. The features were scored individually from 0 to 3, with 0 representing the healthiest state and 3 the maximum degenerative state [43].
Immunochemistry
Aggrecan immunohistochemistry (IHC) was performed on deparaffinized coronal IVD tissue sections of 4-μm thickness. The sections were pretreated with chondroitinase ABC (0.25 U/mL; C3667 MilliporeSigma) to unmask epitopes on aggrecan. Endogenous peroxidase activity was eliminated by treatment with 3% hydrogen peroxide and the sections pre-blocked with a 15 min incubation with Avidin D solution followed by a 15 min incubation with biotin solution (SP-2001, Vector laboratories). Primary antibody against aggrecan (1:200; AB1031, MilliporeSigma) was applied at 4° C overnight, Biotinylated goat anti-rabbit secondary antibody (1:200; BA-1000, Vector Laboratories) was applied for 30 min and the AEC substrate/chromogen KIT (ACG500, ScyTek Laboratories) was used for signal development. All stained slides were mounted with Cytoseal™ 60 (8310-4, Thermo Fisher Scientific) - and were visualized on a Nikon Eclipse E800 microscope [45, 67] and imaged at 100x and 200x. The mean aggrecan signal intensity within NP tissue was quantified by Image J using an established protocol [79].
IVD aggrecan proteolytic degradation by Western blot
Ten caudal intervertebral discs were harvested from each animal, their wet weight recorded and immersed in a 30-fold volume of 4M guanidine hydrochloride solution that was agitated for 48 hours at 4° C to extract the proteoglycan. After centrifugation, the supernatant was collected and the proteoglycans precipitated with 100% ethanol (9:1, v:v). The resulting pellet was then resuspended in ChABC buffer and incubated with 100mU/ml Chondroitinase (Sigma) overnight at 37C. Equal volumes of each sample were mixed with Laemmli buffer (Bio-Rad) and denatured at 95° C for 5 min before electrophoresis. Proteins were fractionated electrophoretically on 4-20% gradient Mini-Protean precast protein gels (Bio-Rad) and then transferred to a polyvinylidene fluoride (PVDF) membrane using a Mini Trans-Blot Blotting System (Bio-Rad). The membrane was blocked with 1% BSA (Bio-Rad), diluted with TBST (0.1% Tween 20 (Sigma-Aldrich) in 1 × Tris-Buffered Saline (TBS, Gibco/Thermo Fisher Scientific) at room temperature for 1 h, washed, and incubated with a primary anti-G1 antibody against aggrecan (rabbit anti-rat aggrecan, Abcam AB36861) at 4° C overnight on a rotating shaker set to 120 rpm at 4° C. Next, the membrane was washed 5 times with TBST buffer and incubated with HRP-conjugated goat anti-rabbit IgG (cat# 31460, Thermo Fisher Scientific/Invitrogen) for 1 h at room temperature. After washing 5 times with TBST, the membrane was incubated with SuperSignal West Pico PLUS Chemiluminescent Substrate. (Cat#: 34577, Thermo Fisher Scientific) Images were acquired using the ChemiDocTM Touch Imaging System (Bio-Rad). Relative protein levels of MMP- and ADAMTS- generated aggrecan fragments were quantitated using ImageLab software.
Tissue RNA isolation and real-time RT-PCR analysis
Using a dissecting microscope, whole intervertebral discs (NP and AF) were dissected from lumbar (L1-L3) and caudal discs (C1-C5) of each mouse in all six cohorts. Pooled tissues from a single animal served as an individual sample. Samples were homogenized, and total RNA was extracted using the RNeasy Plus Universal Mini kit (Qiagen) according to manufacturer’s instruction. A two-step RT-PCR method was employed. Purified, DNA-free RNA was first converted to cDNA using the Superscript IV VILO Master Mix protocol (Invitrogen). Template cDNA (8 ng) and gene-specific primers (IDT, IN) were added to PowerTrack SYBR Green master mix, and expression was quantified using the Step One Plus Real-time PCR System (Applied Biosystems). Relative gene expression was calculated using the ΔΔCt method, with gene expression levels normalized to the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The sequences of primers for each gene are listed in Table 1.
Table 1. Primer Sequences used for RT-PCR.
| Gene | Forward primer (5’-3’) | Reverse primer (5’-3’) |
| GAPDH | GAGGCCGGTGCTGAGTAT | GCGGAGATGATGACCCTTTTGG |
| p16INK4 | AATCTCCGCGAGGAAAGC | GTCTGCAGCGGACTCCAT |
| IL6 | GACTTCCATCCAGTTGCCTTC | ATTTCCACGATTTCCCAGAG |
| IL8 | TCATCATGCTGTTCTGCTACGGG | TCAGCATGGTTCTCATGAGGGTG |
| TNFα | AACAGAAACTCCAGAACATCTTGG | GTCGCGGATCATGCTTTCTG |
Statistics and reproducibility
All statistical analyses were performed using Prism7 (GraphPad). Data are represented as mean ± SEM. Data distribution was assessed with the Shapiro-Wilk normality test, and the differences between the two groups were analyzed by t-test or Mann–Whitney, as appropriate. The differences between three groups were analyzed by ANOVA or Kruskal–Wallis for non-normally distributed data, followed by a Dunn’s multiple comparison test. A χ2-test was used to analyze the differences between the distribution of percentages. p ≤ 0.05 was considered a statistically significant difference.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
PROTAC: Proteolysis Targeting Chimera; BCL: B-Cell Lymphoma (family of proteins); SnC: Senescent Cell; IDD: Intervertebral Disc Degeneration; IVD: Intervertebral Disc; MMP: Matrix Metalloproteinase; IL: Interleukin; TNF: Tumor Necrosis Factor; LBP: Low Back Pain; SASP: Senescence-Associated Secretory Phenotype; ADAMTS: A Disintegrin and Metalloproteinase with Thrombospondin Motifs; SCAP: Senescent Cell Anti-Apoptotic Pathway; MAPK: Mitogen-Activated Protein Kinase; mTOR: Mechanistic Target of Rapamycin; D+Q: Dasatinib and Quercetin; PLGA: Poly(lactic-co-glycolic acid); VHL: von Hippel-Lindau (factor); IP: Intraperitoneal; MERCY: M: Morphological changes in tissue structure; E: Extent of inflammation present in the sample; R: Regeneration potential of the tissue; C: Cellular necrosis or damage observed; Y: Yield of specific histological features relevant to diagnosis (for histological scoring); AF: Annulus Fibrosus; NP: Nucleus Pulposus; EP: Endplate; IGD: Interglobular Domain; GAG: Glycosaminoglycan; RT-PCR: Real-Time Polymerase Chain Reaction; NFκB: Nuclear factor (kappa B); PG: Proteoglycan; 17aE2: 17 alpha estradiol; NDGA: Nordihydroguaiaretic Acid; PEG: Polyethylene Glycol; PHOSAL: Phosphoric Acid Salts; H&E: Hematoxylin and Eosin; IHC: Immunohistochemistry; PVDF: Polyvinylidene Fluoride; BSA: Bovine Serum Albumin; TBST: Tris-Buffered Saline with Triton-X100; L: Lumbar (as in L1-L3); C: Caudal (as in C1-C5); RNA: Ribonucleic Acid; cDNA: Complementary Deoxyribonucleic Acid; PCR: Polymerase Chain Reaction; GAPDH: Glyceraldehyde 3-Phosphate Dehydrogenase; SEM: Standard Error of the Mean; ANOVA: Analysis of Variance.
Author Contributions
Study was conceptualized by Daohong Zhou and Nam V Vo. The animal model, treatment, husbandry and tissue distribution were performed by Bowen Yan and Yang Yang. 753b was synthesized and provided by Peiyi Zhang and Guangrong Zheng. Material preparation, data collection and analysis were performed by Dong Wang, Qing Dong, Karen L. Clark, Ting Zhang and Peter G. Alexander. The first draft of the manuscript was written by Peter G Alexander and all authors commented on previous versions of the manuscript. The Funding was acquired by Daohong Zhou, Guangrong Zheng, Nam V Vo, Gwendolyn A Sowa, Joon Y Lee, and Paul D Robbins. All authors read and approved the final manuscript.
Acknowledgements
We would like to thank the Ferguson Laboratories for Orthopaedic Research, the Bethel Musculoskeletal Research Center and the Department of Orthopaedic Surgery for their support.
Conflicts of Interest
DZ, GZ, PZ, and YY are inventors for the patent applications for use of BCL-XL/2 PROTACs as senolytic and anti-tumor agents. DZ and GZ are co-founders of and have equity in Dialectic Therapeutics, which develops BCL-XL PROTACs to treat cancer. All other authors have nothing to declare. The authors declare that the manuscript in part or in full has not been submitted or published anywhere and will not be submitted elsewhere until the editorial process is completed.
Ethical Statement
All animal procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of The University of Florida and were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The approved protocol number is #202210627.
Funding
This work was supported by the National Institutes of Health grants R01AG081293 (NVV, PDA, JYL), Bethel Musculoskeletal Research Center (NVV, PDA, JYL), R01AG063801 (DZ, GZ), R01CA211963 (DZ, GZ), R01 AG069819 (PDR), P01 AG043376 (PDR), U19 AG056278 (PDR), P01 AG062413 (PDR), U54 AG079754 (PDR), P01 AI172501 (PDR) and U54 AG076041 (PDR), The Orland Endowed Professorship (JYL), and the Department of Orthopaedic Surgery at the University of Pittsburgh.
References
- 1. Davies C, Nitz AJ, Mattacola CG, Kitzman P, Howell D, Viele K, Baxter D, Brockopp D. Practice patterns when treating patients with low back pain: a survey of physical therapists. Physiother Theory Pract. 2014; 30:399–408. https://doi.org/10.3109/09593985.2013.877547 [PubMed]
- 2. Manchikanti L, Singh V, Falco FJ, Benyamin RM, Hirsch JA. Epidemiology of low back pain in adults. Neuromodulation. 2014; 17:3–10. https://doi.org/10.1111/ner.12018 [PubMed]
- 3. Kerr GJ, Veras MA, Kim MK, Séguin CA. Decoding the intervertebral disc: Unravelling the complexities of cell phenotypes and pathways associated with degeneration and mechanotransduction. Semin Cell Dev Biol. 2017; 62:94. https://doi.org/10.1016/j.semcdb.2016.05.008 [PubMed]
- 4. Kos N, Gradisnik L, Velnar T. A Brief Review of the Degenerative Intervertebral Disc Disease. Med Arch. 2019; 73:421–4. https://doi.org/10.5455/medarh.2019.73.421-424 [PubMed]
- 5. Silwal P, Nguyen-Thai AM, Mohammad HA, Wang Y, Robbins PD, Lee JY, Vo NV. Cellular Senescence in Intervertebral Disc Aging and Degeneration: Molecular Mechanisms and Potential Therapeutic Opportunities. Biomolecules. 2023; 13:686. https://doi.org/10.3390/biom13040686 [PubMed]
- 6. Hoffeld K, Lenz M, Egenolf P, Weber M, Heck V, Eysel P, Scheyerer MJ. Patient-related risk factors and lifestyle factors for lumbar degenerative disc disease: a systematic review. Neurochirurgie. 2023; 69:101482. https://doi.org/10.1016/j.neuchi.2023.101482 [PubMed]
- 7. Parenteau CS, Lau EC, Campbell IC, Courtney A. Prevalence of spine degeneration diagnosis by type, age, gender, and obesity using Medicare data. Sci Rep. 2021; 11:5389. https://doi.org/10.1038/s41598-021-84724-6 [PubMed]
- 8. Vo NV, Hartman RA, Patil PR, Risbud MV, Kletsas D, Iatridis JC, Hoyland JA, Le Maitre CL, Sowa GA, Kang JD. Molecular mechanisms of biological aging in intervertebral discs. J Orthop Res. 2016; 34:1289–306. https://doi.org/10.1002/jor.23195 [PubMed]
- 9. Wang F, Cai F, Shi R, Wang XH, Wu XT. Aging and age related stresses: a senescence mechanism of intervertebral disc degeneration. Osteoarthritis Cartilage. 2016; 24:398–408. https://doi.org/10.1016/j.joca.2015.09.019 [PubMed]
- 10. Herdy JR, Traxler L, Agarwal RK, Karbacher L, Schlachetzki JC, Boehnke L, Zangwill D, Galasko D, Glass CK, Mertens J, Gage FH. Increased post-mitotic senescence in aged human neurons is a pathological feature of Alzheimer's disease. Cell Stem Cell. 2022; 29:1637–52.e6. https://doi.org/10.1016/j.stem.2022.11.010 [PubMed]
- 11. Walaszczyk A, Dookun E, Redgrave R, Tual-Chalot S, Victorelli S, Spyridopoulos I, Owens A, Arthur HM, Passos JF, Richardson GD. Pharmacological clearance of senescent cells improves survival and recovery in aged mice following acute myocardial infarction. Aging Cell. 2019; 18:e12945. https://doi.org/10.1111/acel.12945 [PubMed]
- 12. Hickson LJ, Langhi Prata LG, Bobart SA, Evans TK, Giorgadze N, Hashmi SK, Herrmann SM, Jensen MD, Jia Q, Jordan KL, Kellogg TA, Khosla S, Koerber DM, et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine. 2019; 52:102595. https://doi.org/10.1016/j.ebiom.2019.12.004 [PubMed]. Erratum in: EBioMedicine. 2019; 47:446–56. DOI:10.1016/j.ebiom.2019.08.069 PMID:31542391.
- 13. Kim H, Jang J, Song MJ, Kim G, Park CH, Lee DH, Lee SH, Chung JH. Attenuation of intrinsic ageing of the skin via elimination of senescent dermal fibroblasts with senolytic drugs. J Eur Acad Dermatol Venereol. 2022; 36:1125–35. https://doi.org/10.1111/jdv.18051 [PubMed]
- 14. Dungan CM, Murach KA, Zdunek CJ, Tang ZJ, Nolt GL, Brightwell CR, Hettinger Z, Englund DA, Liu Z, Fry CS, Filareto A, Franti M, Peterson CA. Deletion of SA β-Gal+ cells using senolytics improves muscle regeneration in old mice. Aging Cell. 2022; 21:e13528. https://doi.org/10.1111/acel.13528 [PubMed]
- 15. Miura Y, Endo K, Komori K, Sekiya I. Clearance of senescent cells with ABT-263 improves biological functions of synovial mesenchymal stem cells from osteoarthritis patients. Stem Cell Res Ther. 2022; 13:222. https://doi.org/10.1186/s13287-022-02901-4 [PubMed]
- 16. Farr JN, Khosla S. Cellular senescence in bone. Bone. 2019; 121:121–33. https://doi.org/10.1016/j.bone.2019.01.015 [PubMed]
- 17. Kaur J, Farr JN. Cellular senescence in age-related disorders. Transl Res. 2020; 226:96–104. https://doi.org/10.1016/j.trsl.2020.06.007 [PubMed]
- 18. Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: senolytics and senomorphics. FEBS J. 2023; 290:1362–83. https://doi.org/10.1111/febs.16350 [PubMed]
- 19. Novais EJ, Narayanan R, Canseco JA, van de Wetering K, Kepler CK, Hilibrand AS, Vaccaro AR, Risbud MV. A new perspective on intervertebral disc calcification-from bench to bedside. Bone Res. 2024; 12:3. https://doi.org/10.1038/s41413-023-00307-3 [PubMed]
- 20. Zhao Z, Wang Y, Wang Z, Zhang F, Ding Z, Fan T. Senescence in Intervertebral Disc Degeneration: A Comprehensive Analysis Based on Bioinformatic Strategies. Immun Inflamm Dis. 2024; 12:e70072. https://doi.org/10.1002/iid3.70072 [PubMed]
- 21. Sun Q, Tian FM, Liu F, Fang JK, Hu YP, Lian QQ, Zhou Z, Zhang L. Denosumab alleviates intervertebral disc degeneration adjacent to lumbar fusion by inhibiting endplate osteochondral remodeling and vertebral osteoporosis in ovariectomized rats. Arthritis Res Ther. 2021; 23:152. https://doi.org/10.1186/s13075-021-02525-8 [PubMed]
- 22. Khosla S, Farr JN, Tchkonia T, Kirkland JL. The role of cellular senescence in ageing and endocrine disease. Nat Rev Endocrinol. 2020; 16:263–75. https://doi.org/10.1038/s41574-020-0335-y [PubMed]
- 23. Abildgaard J, Tingstedt J, Zhao Y, Hartling HJ, Pedersen AT, Lindegaard B, Dam Nielsen S. Increased systemic inflammation and altered distribution of T-cell subsets in postmenopausal women. PLoS One. 2020; 15:e0235174. https://doi.org/10.1371/journal.pone.0235174 [PubMed]
- 24. Zhou L, Li C, Zhang H. Correlation between bone mineral density of different sites and lumbar disc degeneration in postmenopausal women. Medicine (Baltimore). 2022; 101:e28947. https://doi.org/10.1097/MD.0000000000028947 [PubMed]
- 25. Novais EJ, Tran VA, Johnston SN, Darris KR, Roupas AJ, Sessions GA, Shapiro IM, Diekman BO, Risbud MV. Long-term treatment with senolytic drugs Dasatinib and Quercetin ameliorates age-dependent intervertebral disc degeneration in mice. Nat Commun. 2021; 12:5213. https://doi.org/10.1038/s41467-021-25453-2 [PubMed]
- 26. Gasek NS, Kuchel GA, Kirkland JL, Xu M. Strategies for Targeting Senescent Cells in Human Disease. Nat Aging. 2021; 1:870–9. https://doi.org/10.1038/s43587-021-00121-8 [PubMed]
- 27. Hu L, Li H, Zi M, Li W, Liu J, Yang Y, Zhou D, Kong QP, Zhang Y, He Y. Why Senescent Cells Are Resistant to Apoptosis: An Insight for Senolytic Development. Front Cell Dev Biol. 2022; 10:822816. https://doi.org/10.3389/fcell.2022.822816 [PubMed]
- 28. Tchkonia T, Palmer AK, Kirkland JL. New Horizons: Novel Approaches to Enhance Healthspan Through Targeting Cellular Senescence and Related Aging Mechanisms. J Clin Endocrinol Metab. 2021; 106:e1481–7. https://doi.org/10.1210/clinem/dgaa728 [PubMed]
- 29. Cherif H, Bisson DG, Mannarino M, Rabau O, Ouellet JA, Haglund L. Senotherapeutic drugs for human intervertebral disc degeneration and low back pain. Elife. 2020; 9:e54693. https://doi.org/10.7554/eLife.54693 [PubMed]
- 30. Lim S, An SB, Jung M, Joshi HP, Kumar H, Kim C, Song SY, Lee JR, Kang M, Han I, Kim BS. Local Delivery of Senolytic Drug Inhibits Intervertebral Disc Degeneration and Restores Intervertebral Disc Structure. Adv Healthc Mater. 2022; 11:e2101483. https://doi.org/10.1002/adhm.202101483 [PubMed]
- 31. Negi A, Voisin-Chiret AS. Strategies to Reduce the On-Target Platelet Toxicity of Bcl-xL Inhibitors: PROTACs, SNIPERs and Prodrug-Based Approaches. Chembiochem. 2022; 23:e202100689. https://doi.org/10.1002/cbic.202100689 [PubMed]
- 32. Lv D, Pal P, Liu X, Jia Y, Thummuri D, Zhang P, Hu W, Pei J, Zhang Q, Zhou S, Khan S, Zhang X, Hua N, et al. Development of a BCL-xL and BCL-2 dual degrader with improved anti-leukemic activity. Nat Commun. 2021; 12:6896. https://doi.org/10.1038/s41467-021-27210-x [PubMed]
- 33. Jia Y, Han L, Ramage CL, Wang Z, Weng CC, Yang L, Colla S, Ma H, Zhang W, Andreeff M, Daver N, Jain N, Pemmaraju N, et al. Co-targeting BCL-XL and BCL-2 by PROTAC 753B eliminates leukemia cells and enhances efficacy of chemotherapy by targeting senescent cells. Haematologica. 2023; 108:2626–38. https://doi.org/10.3324/haematol.2022.281915 [PubMed]
- 34. Khan S, Cao L, Wiegand J, Zhang P, Zajac-Kaye M, Kaye FJ, Zheng G, Zhou D. PROTAC-Mediated Dual Degradation of BCL-xL and BCL-2 Is a Highly Effective Therapeutic Strategy in Small-Cell Lung Cancer. Cells. 2024; 13:528. https://doi.org/10.3390/cells13060528 [PubMed]
- 35. Yang Y, Jn-Simon N, He Y, Sun C, Zhang P, Hu W, Tian T, Zeng H, Basha S, Huerta AS, Sun LZ, Yin XM, Hromas R, et al. A BCL-xL/BCL-2 PROTAC effectively clears senescent cells in the liver and reduces MASH-driven hepatocellular carcinoma in mice. Nat Aging. 2025; 5:386–400. https://doi.org/10.1038/s43587-025-00811-7 [PubMed]
- 36. Gullbrand SE, Peterson J, Mastropolo R, Roberts TT, Lawrence JP, Glennon JC, DiRisio DJ, Ledet EH. Low rate loading-induced convection enhances net transport into the intervertebral disc in vivo. Spine J. 2015; 15:1028–33. https://doi.org/10.1016/j.spinee.2014.12.003 [PubMed]
- 37. Karppinen J, Inkinen RI, Kääpä E, Lammi MJ, Tammi MI, Holm S, Vanharanta H. Effects of tiaprofenic acid and indomethacin on proteoglycans in the degenerating porcine intervertebral disc. Spine (Phila Pa 1976). 1995; 20:1170–7. https://doi.org/10.1097/00007632-199505150-00012 [PubMed]
- 38. Aliyev A, Saboury B, Kwee TC, Torigian DA, Basu S, Wulff Christensen H, Alavi A. Age-related inflammatory changes in the spine as demonstrated by (18)F-FDG-PET:observation and insight into degenerative spinal changes. Hell J Nucl Med. 2012; 15:197–201. https://doi.org/10.1967/s002449910054 [PubMed]
- 39. Chen CH, Chiang CJ, Wu LC, Yang CH, Kuo YJ, Tsai TH. In vitro Penetration and in vivo Distribution of Honokiol into the Intervertebral Disc in Rat. Anal Sci. 2015; 31:1297–302. https://doi.org/10.2116/analsci.31.1297 [PubMed]
- 40. Tseng HW, Glant TT, Brown MA, Kenna TJ, Thomas GP, Pettit AR. Early anti-inflammatory intervention ameliorates axial disease in the proteoglycan-induced spondylitis mouse model of ankylosing spondylitis. BMC Musculoskelet Disord. 2017; 18:228. https://doi.org/10.1186/s12891-017-1600-7 [PubMed]
- 41. Hanberg P, Bue M, Jørgensen AR, Thomassen M, Öbrink-Hansen K, Søballe K, Stilling M. Pharmacokinetics of double-dose cefuroxime in porcine intervertebral disc and vertebral cancellous bone-a randomized microdialysis study. Spine J. 2020; 20:1327–32. https://doi.org/10.1016/j.spinee.2020.03.006 [PubMed]
- 42. Walters R, Moore R, Fraser R. Penetration of cephazolin in human lumbar intervertebral disc. Spine (Phila Pa 1976). 2006; 31:567–70. https://doi.org/10.1097/01.brs.0000201244.24003.2d [PubMed]
- 43. Melgoza IP, Chenna SS, Tessier S, Zhang Y, Tang SY, Ohnishi T, Novais EJ, Kerr GJ, Mohanty S, Tam V, Chan WC, Zhou CM, Zhang Y, et al. Development of a standardized histopathology scoring system using machine learning algorithms for intervertebral disc degeneration in the mouse model-An ORS spine section initiative. JOR Spine. 2021; 4:e1164. https://doi.org/10.1002/jsp2.1164 [PubMed]
- 44. Roughley PJ, Alini M, Antoniou J. The role of proteoglycans in aging, degeneration and repair of the intervertebral disc. Biochem Soc Trans. 2002; 30:869–74. https://doi.org/10.1042/bst0300869 [PubMed]
- 45. Patil P, Dong Q, Wang D, Chang J, Wiley C, Demaria M, Lee J, Kang J, Niedernhofer LJ, Robbins PD, Sowa G, Campisi J, Zhou D, Vo N. Systemic clearance of p16INK4a -positive senescent cells mitigates age-associated intervertebral disc degeneration. Aging Cell. 2019; 18:e12927. https://doi.org/10.1111/acel.12927 [PubMed]
- 46. Le Maitre CL, Freemont AJ, Hoyland JA. Accelerated cellular senescence in degenerate intervertebral discs: a possible role in the pathogenesis of intervertebral disc degeneration. Arthritis Res Ther. 2007; 9:R45. https://doi.org/10.1186/ar2198 [PubMed]
- 47. Zhong J, Chen J, Oyekan AA, Epperly MW, Greenberger JS, Lee JY, Sowa GA, Vo NV. Ionizing Radiation Induces Disc Annulus Fibrosus Senescence and Matrix Catabolism via MMP-Mediated Pathways. Int J Mol Sci. 2022; 23:4014. https://doi.org/10.3390/ijms23074014 [PubMed]
- 48. Nasto LA, Wang D, Robinson AR, Clauson CL, Ngo K, Dong Q, Roughley P, Epperly M, Huq SM, Pola E, Sowa G, Robbins PD, Kang J, et al. Genotoxic stress accelerates age-associated degenerative changes in intervertebral discs. Mech Ageing Dev. 2013; 134:35–42. https://doi.org/10.1016/j.mad.2012.11.002 [PubMed]
- 49. Ngo K, Patil P, McGowan SJ, Niedernhofer LJ, Robbins PD, Kang J, Sowa G, Vo N. Senescent intervertebral disc cells exhibit perturbed matrix homeostasis phenotype. Mech Ageing Dev. 2017; 166:16–23. https://doi.org/10.1016/j.mad.2017.08.007 [PubMed]
- 50. Zhu Y, Tchkonia T, Pirtskhalava T, Gower AC, Ding H, Giorgadze N, Palmer AK, Ikeno Y, Hubbard GB, Lenburg M, O'Hara SP, LaRusso NF, Miller JD, et al. The Achilles' heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell. 2015; 14:644–58. https://doi.org/10.1111/acel.12344 [PubMed]
- 51. Novais EJ, Ottone OK, Jagannath S, Akande EJ, Barve RA, Risbud MV. Dasatinib and quercetin senolytic treatment delays early onset intervertebral disc degeneration in SM/J mice. Bone Res. 2026; 14:42. https://doi.org/10.1038/s41413-026-00526-4 [PubMed]
- 52. Vo NV, Hartman RA, Yurube T, Jacobs LJ, Sowa GA, Kang JD. Expression and regulation of metalloproteinases and their inhibitors in intervertebral disc aging and degeneration. Spine J. 2013; 13:331–41. https://doi.org/10.1016/j.spinee.2012.02.027 [PubMed]
- 53. Han EH, Chen SS, Klisch SM, Sah RL. Contribution of proteoglycan osmotic swelling pressure to the compressive properties of articular cartilage. Biophys J. 2011; 101:916–24. https://doi.org/10.1016/j.bpj.2011.07.006 [PubMed]
- 54. Iatridis JC, MacLean JJ, O'Brien M, Stokes IA. Measurements of proteoglycan and water content distribution in human lumbar intervertebral discs. Spine (Phila Pa 1976). 2007; 32:1493–7. https://doi.org/10.1097/BRS.0b013e318067dd3f [PubMed]
- 55. Ruffilli A, Viroli G, Neri S, Traversari M, Barile F, Manzetti M, Assirelli E, Ialuna M, Vita F, Faldini C. Mechanobiology of the Human Intervertebral Disc: Systematic Review of the Literature and Future Perspectives. Int J Mol Sci. 2023; 24:2728. https://doi.org/10.3390/ijms24032728 [PubMed]
- 56. Wang D, He X, Wang D, Peng P, Xu X, Gao B, Zheng C, Wang H, Jia H, Shang Q, Sun Z, Luo Z, Yang L. Quercetin Suppresses Apoptosis and Attenuates Intervertebral Disc Degeneration via the SIRT1-Autophagy Pathway. Front Cell Dev Biol. 2020; 8:613006. https://doi.org/10.3389/fcell.2020.613006 [PubMed]
- 57. Zhang S, Liang W, Abulizi Y, Xu T, Cao R, Xun C, Zhang J, Sheng W. Quercetin Alleviates Intervertebral Disc Degeneration by Modulating p38 MAPK-Mediated Autophagy. Biomed Res Int. 2021; 2021:6631562. https://doi.org/10.1155/2021/6631562 [PubMed]
- 58. Zhao WJ, Liu X, Hu M, Zhang Y, Shi PZ, Wang JW, Lu XH, Cheng XF, Tao YP, Feng XM, Wang YX, Zhang L. Quercetin ameliorates oxidative stress-induced senescence in rat nucleus pulposus-derived mesenchymal stem cells via the miR-34a-5p/SIRT1 axis. World J Stem Cells. 2023; 15:842–65. https://doi.org/10.4252/wjsc.v15.i8.842 [PubMed]
- 59. Mannarino M, Cherif H, Li L, Sheng K, Rabau O, Jarzem P, Weber MH, Ouellet JA, Haglund L. Toll-like receptor 2 induced senescence in intervertebral disc cells of patients with back pain can be attenuated by o-vanillin. Arthritis Res Ther. 2021; 23:117. https://doi.org/10.1186/s13075-021-02504-z [PubMed]
- 60. Harrison DE, Strong R, Allison DB, Ames BN, Astle CM, Atamna H, Fernandez E, Flurkey K, Javors MA, Nadon NL, Nelson JF, Pletcher S, Simpkins JW, et al. Acarbose, 17-α-estradiol, and nordihydroguaiaretic acid extend mouse lifespan preferentially in males. Aging Cell. 2014; 13:273–82. https://doi.org/10.1111/acel.12170 [PubMed]
- 61. Strong R, Miller RA, Astle CM, Floyd RA, Flurkey K, Hensley KL, Javors MA, Leeuwenburgh C, Nelson JF, Ongini E, Nadon NL, Warner HR, Harrison DE. Nordihydroguaiaretic acid and aspirin increase lifespan of genetically heterogeneous male mice. Aging Cell. 2008; 7:641–50. https://doi.org/10.1111/j.1474-9726.2008.00414.x [PubMed]
- 62. Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, Miller RA. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009; 460:392–5. https://doi.org/10.1038/nature08221 [PubMed]
- 63. Strong R, Miller RA, Antebi A, Astle CM, Bogue M, Denzel MS, Fernandez E, Flurkey K, Hamilton KL, Lamming DW, Javors MA, de Magalhães JP, Martinez PA, et al. Longer lifespan in male mice treated with a weakly estrogenic agonist, an antioxidant, an α-glucosidase inhibitor or a Nrf2-inducer. Aging Cell. 2016; 15:872–84. https://doi.org/10.1111/acel.12496 [PubMed]
- 64. Kumari R, Jat P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front Cell Dev Biol. 2021; 9:645593. https://doi.org/10.3389/fcell.2021.645593 [PubMed]
- 65. Chen B, Zhu R, Hu H, Zhan M, Wang T, Huang F, Wei F, Chai Y, Ling Z, Zou X. Elimination of Senescent Cells by Senolytics Facilitates Bony Endplate Microvessel Formation and Mitigates Disc Degeneration in Aged Mice. Front Cell Dev Biol. 2022; 10:853688. https://doi.org/10.3389/fcell.2022.853688 [PubMed]
- 66. Chang J, Wang Y, Shao L, Laberge RM, Demaria M, Campisi J, Janakiraman K, Sharpless NE, Ding S, Feng W, Luo Y, Wang X, Aykin-Burns N, et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat Med. 2016; 22:78–83. https://doi.org/10.1038/nm.4010 [PubMed]
- 67. Lei C, Colangelo D, Patil P, Li V, Ngo K, Wang D, Dong Q, Yousefzadeh MJ, Lin H, Lee J, Kang J, Sowa G, Wyss-Coray T, et al. Influences of circulatory factors on intervertebral disc aging phenotype. Aging (Albany NY). 2020; 12:12285–304. https://doi.org/10.18632/aging.103421 [PubMed]
- 68. Gulej R, Nyúl-Tóth Á, Ahire C, DelFavero J, Balasubramanian P, Kiss T, Tarantini S, Benyo Z, Pacher P, Csik B, Yabluchanskiy A, Mukli P, Kuan-Celarier A, et al. Elimination of senescent cells by treatment with Navitoclax/ABT263 reverses whole brain irradiation-induced blood-brain barrier disruption in the mouse brain. Geroscience. 2023; 45:2983–3002. https://doi.org/10.1007/s11357-023-00870-x [PubMed]
- 69. Riley LH 3r, Banovac K, Martinez OV, Eismont FJ. Tissue distribution of antibiotics in the intervertebral disc. Spine (Phila Pa 1976). 1994; 19:2619–25. [PubMed]
- 70. Yang H, Chen C, Chen H, Duan X, Li J, Zhou Y, Zeng W, Yang L. Navitoclax (ABT263) reduces inflammation and promotes chondrogenic phenotype by clearing senescent osteoarthritic chondrocytes in osteoarthritis. Aging (Albany NY). 2020; 12:12750–70. https://doi.org/10.18632/aging.103177 [PubMed]
- 71. Kim HN, Chang J, Shao L, Han L, Iyer S, Manolagas SC, O'Brien CA, Jilka RL, Zhou D, Almeida M. DNA damage and senescence in osteoprogenitors expressing Osx1 may cause their decrease with age. Aging Cell. 2017; 16:693–703. https://doi.org/10.1111/acel.12597 [PubMed]
- 72. Ackert-Bicknell CL, Anderson LC, Sheehan S, Hill WG, Chang B, Churchill GA, Chesler EJ, Korstanje R, Peters LL. Aging Research Using Mouse Models. Curr Protoc Mouse Biol. 2015; 5:95–133. https://doi.org/10.1002/9780470942390.mo140195 [PubMed]
- 73. Flurkey K, Currer JM, Harrison DE. Mouse Models in Aging Research. The Mouse in Biomedical Research. Fox JG, Davisson MT, Quimby FW, Barthold SW, Newcomer CE, Smith AL Editors. 2007. Academic Press: Cambridge, MA. p. 637–72. https://doi.org/10.1016/B978-012369454-6/50074-1
- 74. Alvarez-Garcia O, Matsuzaki T, Olmer M, Masuda K, Lotz MK. Age-related reduction in the expression of FOXO transcription factors and correlations with intervertebral disc degeneration. J Orthop Res. 2017; 35:2682–91. https://doi.org/10.1002/jor.23583 [PubMed]
- 75. Ohnishi T, Sudo H, Tsujimoto T, Iwasaki N. Age-related spontaneous lumbar intervertebral disc degeneration in a mouse model. J Orthop Res. 2018; 36:224–32. https://doi.org/10.1002/jor.23634 [PubMed]
- 76. Mylonas KJ, O'Sullivan ED, Humphries D, Baird DP, Docherty MH, Neely SA, Krimpenfort PJ, Melk A, Schmitt R, Ferreira-Gonzalez S, Forbes SJ, Hughes J, Ferenbach DA. Cellular senescence inhibits renal regeneration after injury in mice, with senolytic treatment promoting repair. Sci Transl Med. 2021; 13:eabb0203. https://doi.org/10.1126/scitranslmed.abb0203 [PubMed]
- 77. Sharma AK, Roberts RL, Benson RD
Jr , Pierce JL, Yu K, Hamrick MW, McGee-Lawrence ME. The Senolytic Drug Navitoclax (ABT-263) Causes Trabecular Bone Loss and Impaired Osteoprogenitor Function in Aged Mice. Front Cell Dev Biol. 2020; 8:354. https://doi.org/10.3389/fcell.2020.00354 [PubMed] - 78. Nasto LA, Seo HY, Robinson AR, Tilstra JS, Clauson CL, Sowa GA, Ngo K, Dong Q, Pola E, Lee JY, Niedernhofer LJ, Kang JD, Robbins PD, Vo NV. ISSLS prize winner: inhibition of NF-κB activity ameliorates age-associated disc degeneration in a mouse model of accelerated aging. Spine (Phila Pa 1976). 2012; 37:1819–25. https://doi.org/10.1097/BRS.0b013e31824ee8f7 [PubMed]
- 79. Crowe AR, Yue W. Semi-quantitative Determination of Protein Expression using Immunohistochemistry Staining and Analysis: An Integrated Protocol. Bio Protoc. 2019; 9:e3465. https://doi.org/10.21769/BioProtoc.3465 [PubMed]