



Introduction
Due to the evidently better variability in immunogenicity during tumor progression, melanoma is broadly considered as an immunogenic malignancy [1], which serves as the model system for evaluating the effectiveness of invented immunotherapies [2]. Therefore, therapies for melanoma have been recently changed owing to the emergence of immune checkpoint inhibitors (ICI) including anti-CTLA-4 and anti-PD-1 agents [3]. ICI therapies have observably extended the survival time for advanced melanoma patients [4, 5]. Nevertheless, the remarkable efficacy was only observed in a fraction of patients, most were not benefitted.
Recent studies demonstrated that ICI therapies were influenced by a combination of predictive biomarkers related to genomics, immune checkpoints expression, characteristics of the microenvironment, and gut microbiome [6]. Tumor mutation burden (TMB) and neoantigen burden (NB) emerged as promising markers for evaluating ICI efficacy and previous findings have demonstrated their positive association with the immunotherapy response rate and prognosis via numerous clinical trials [7–9]. However, a few studies concluded controversial results, that is high TMB sometimes could not accurately predict ICI response [8]. Immune checkpoints, such as programmed cell death ligand 1 (PD-L1) expression is another widely used biomarker associated with ICI therapies efficacy. Similarly, it may be out of work in some trials [10]. In view of the current situation, novel and more effective indicators were needed to distinguish subpopulations that are likely to be sensitive to ICI treatment.
Nod-like receptor protein 3 (NLRP3) inflammasome was a three-domain complex associated with inflammation regulation, immune response, and cell apoptosis [11]. Functions of NLRP3 inflammasome in cancer progression remained inconsistent owing to the controversial results reported [12]. For instance, the protective effects of this inflammasome were observed in colon cancer [13, 14]. Conversely, it exhibited a promotion role in cancers of gastric [15], liver [16], head and neck [17], lung [18], prostate [19], glioblastoma [20], and melanoma [21].
Recent studies demonstrated that NLRP3 inflammasome upregulation may inhibit the inflammatory responses in melanoma. Consistently, a mice model with NLRP3 deficiency showed the protection roles against cancer progression [22, 23]. The progression of cancer cells could be suppressed by reduced NLRP3 inflammasome and IL-1β expression [24]. It has been shown that NLRP3 downregulation and reduced IL-1β secretion decreased metastatic melanoma by thymoquinone therapy in a mouse model [21]. Evaluation of the roles of NLRP3 inflammasome in the immune response by employing vaccination against the melanoma cells demonstrated that mice with NLRP3 vaccination deficiency who received a subcutaneous injection of poorly immunogenic melanoma cells leading to a 4-fold promotion in survival times as compared to the control mice [25]. NLRP3 plays vital roles in melanoma tumorigenesis, progression, and immune response, however, its alterations association with ICI efficacy remains unclear.
Herein, we analyzed whether NLRP3 mutations were correlated with immunological and genomic features with publicly available data in melanoma. Further, the association of NLRP3 mutations with ICI efficacy was estimated with an aggregated ICI-treated cohort. Novel findings would provide implications for tailoring clinical trials and immunotherapeutic strategies for melanoma.
Results
NLRP3 mutations in melanoma
In the TCGA cohort, 89 (19.1%) of 467 melanoma patients harbored NLRP3 mutations. NLRP3 is one frequently mutated gene and we found that patients with NLRP3 mutations had higher TML as compared with others (Figure 1A). Of 89 NLRP3 mutated patients, 53 (59.5%) also had mutations of genes related to genomic maintenance including TP53, BRCA1/2, POLE, and MMR genes (Figure 1B). Further analyses revealed that NLRP3 mutated patients harbored significantly higher mutation rates of above genome repair genes than NLRP3 wild-type patients (Fisher exact test, all P < 0.05; Figure 1C). Mutational patterns of NLRP3, its family members, and genomic maintenance genes were exhibited in Figure 1B.
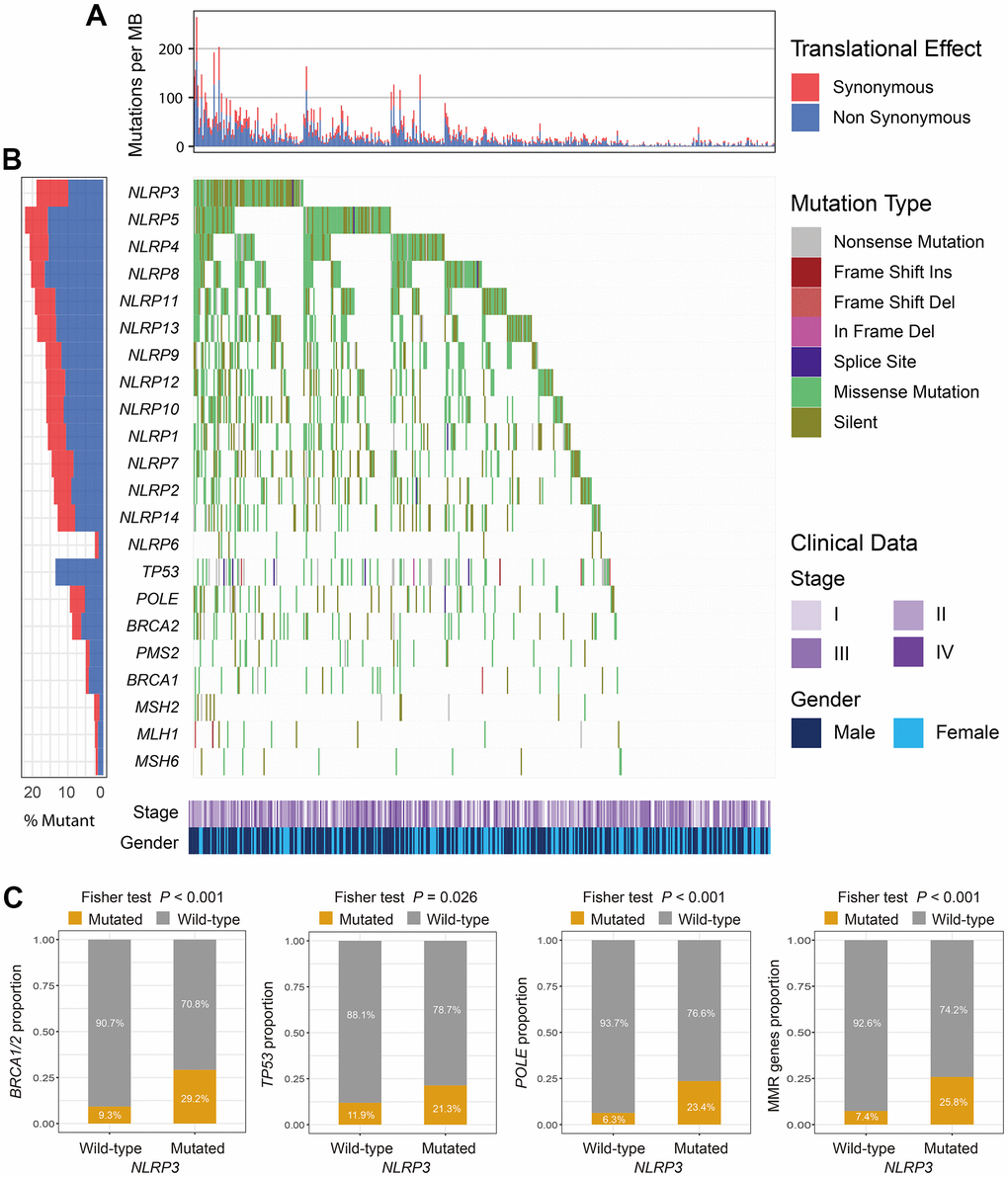
Figure 1. The mutational patterns of NLR family members and genome maintenance genes. (A) TMB stratified by synonymous and non-synonymous mutations for each patient. (B) Waterfall plot for NLR family members and genome maintenance genes. (C) Association of NLRP3 mutations with BRCA1/2, TP53, POLE, and MMR genes mutations.
NLRP3 mutations were correlated with high TMB, NB, and favorable genomic features
In the TCGA cohort, patients with NLRP3 mutations had significantly higher TMB than patients without (Wilcoxon rank-sum test, P < 0.001; Figure 2A). We observed that genome repair regulators including BRCA1/2, TP53, POLE, and MMR genes were frequently mutated and mutations of these genes also induced significantly higher TMB (all P < 0.01; Figure 2B). Multivariate Logistic regression model included clinical variables (i.e., age, gender, and stage) and mutations of above genome repair genes was performed to adjust confounding factors. Association of NLRP3 mutations with higher TMB was still significant (OR: 7.50, 95% CI: 3.85-15.24, P < 0.001; Figure 2B). In 2 independent cohorts from ICGC, the Wilcoxon rank-sum test showed that NLRP3 mutated patients also had significantly higher TMB than NLRP3 wild-type patients (both P < 0.001; Supplementary Figure 1A, 1C). Further multivariate regression model obtained consistent findings after adjusting clinical and genomic confounders (MELA-AU cohort [OR: 8.87, 95% CI: 3.04-29.63, P < 0.001]; SKCA-BR cohort [OR: 10.38, 95% CI: 1.38-101.34, P = 0.028]; Supplementary Figure 1B, 1D).
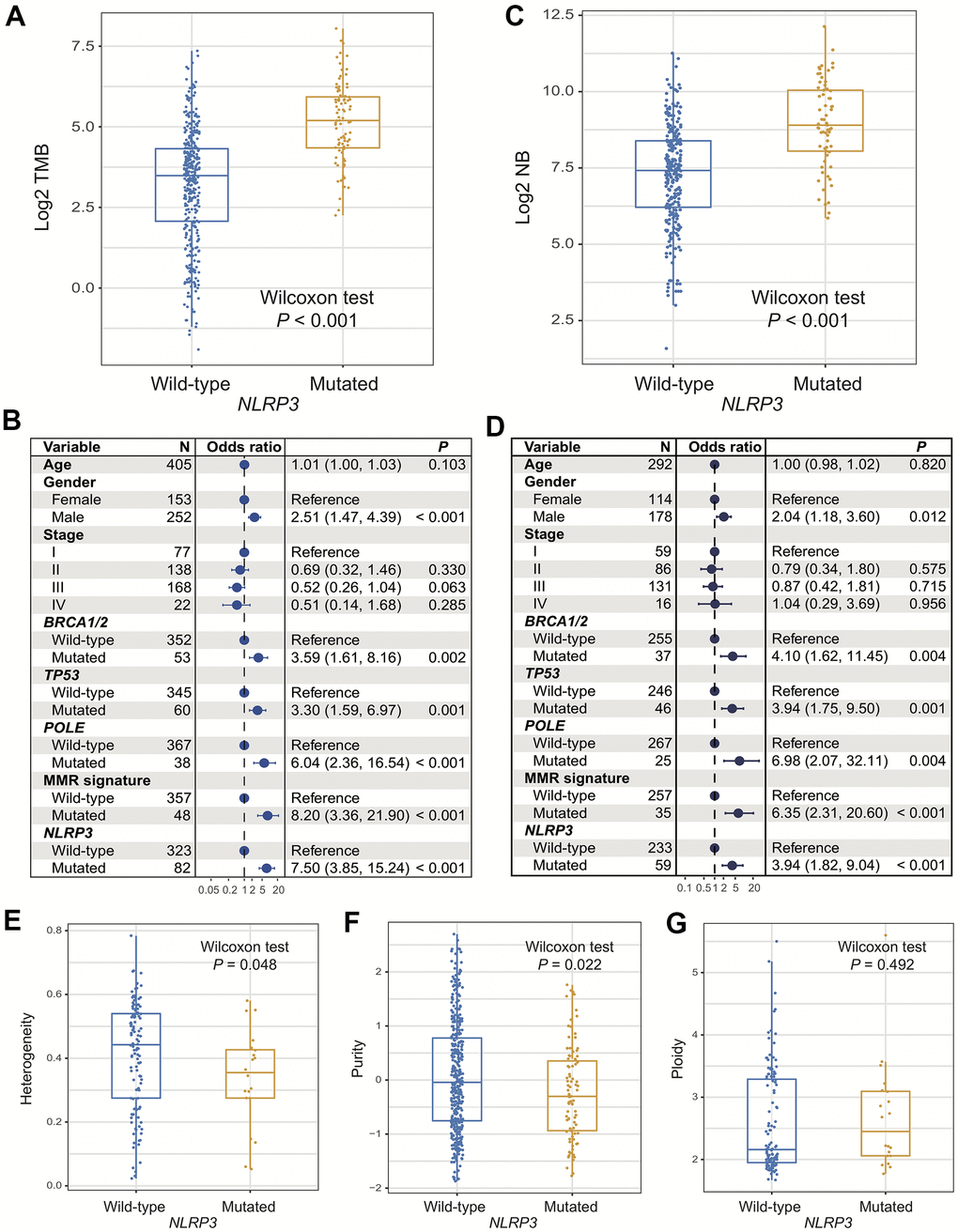
Figure 2. Association of NLRP3 mutations with TMB, NB, and genomic features. (A, B) NLRP3 mutations versus TMB with univariate analysis and multivariate regression model. (C, D) NLRP3 mutations versus NB with univariate analysis and multivariate regression model. NLRP3 mutations association with (E) tumor heterogeneity, (F) purity, and (G) ploidy.
Significantly elevated NB was observed in patients with NLRP3 mutations (Wilcoxon rank-sum test, P < 0.001; Figure 2C). Multivariate Logistic model with other confounders taken into account remained statistically significant (OR: 3.94, 95% CI: 1.82-9.04, P < 0.001; Figure 2D).
Lower tumor heterogeneity and purity, which suggest the lower proportions of subclonal mutations and tumor cells in microenvironment, were statistically associated with NLRP3 mutations (Wilcoxon rank-sum test, P = 0.048 and P = 0.022; Figure 2E, 2F). No significant difference was found in tumor ploidy based on NLRP3 mutational statuses (Wilcoxon rank-sum test, P = 0.49; Figure 2G). The above findings demonstrated that NLRP3 mutations were linked with favorable genomic characteristics.
NLRP3 mutations were correlated with the better microenvironment
ESTIMATE algorithm showed that the difference of overall immune cells infiltration was not significant between NLRP3 mutated and wild-type patients (Wilcoxon rank-sum test, P = 0.329; Supplementary Figure 2). We thus estimated the abundance of distinct immune cell subtypes using the CIBERSORT approach and compared their differences based on NLRP3 statuses. Results revealed that significantly lower enrichment of regulatory T cells and higher enrichment of naive B cells were observed in patients with NLRP3 mutations (Wilcoxon rank-sum test, P = 0.023 and P = 0.039; Figure 3A). We further calculated the distinct immune cells infiltration according to NLRP3 mutational statuses via the Angelova et al. method. Consistent with the result of CIBERSORT, significantly lower infiltration of regulatory T cells was also found in patients with NLRP3 mutations (Wilcoxon rank-sum test, P = 0.045; Supplementary Figure 3). In addition, we observed a higher abundance of activated CD4 T cells, effector CD4 T cells, and dendritic cells in NLRP3 mutant patients (Wilcoxon rank-sum test, all P < 0.05; Supplementary Figure 3).
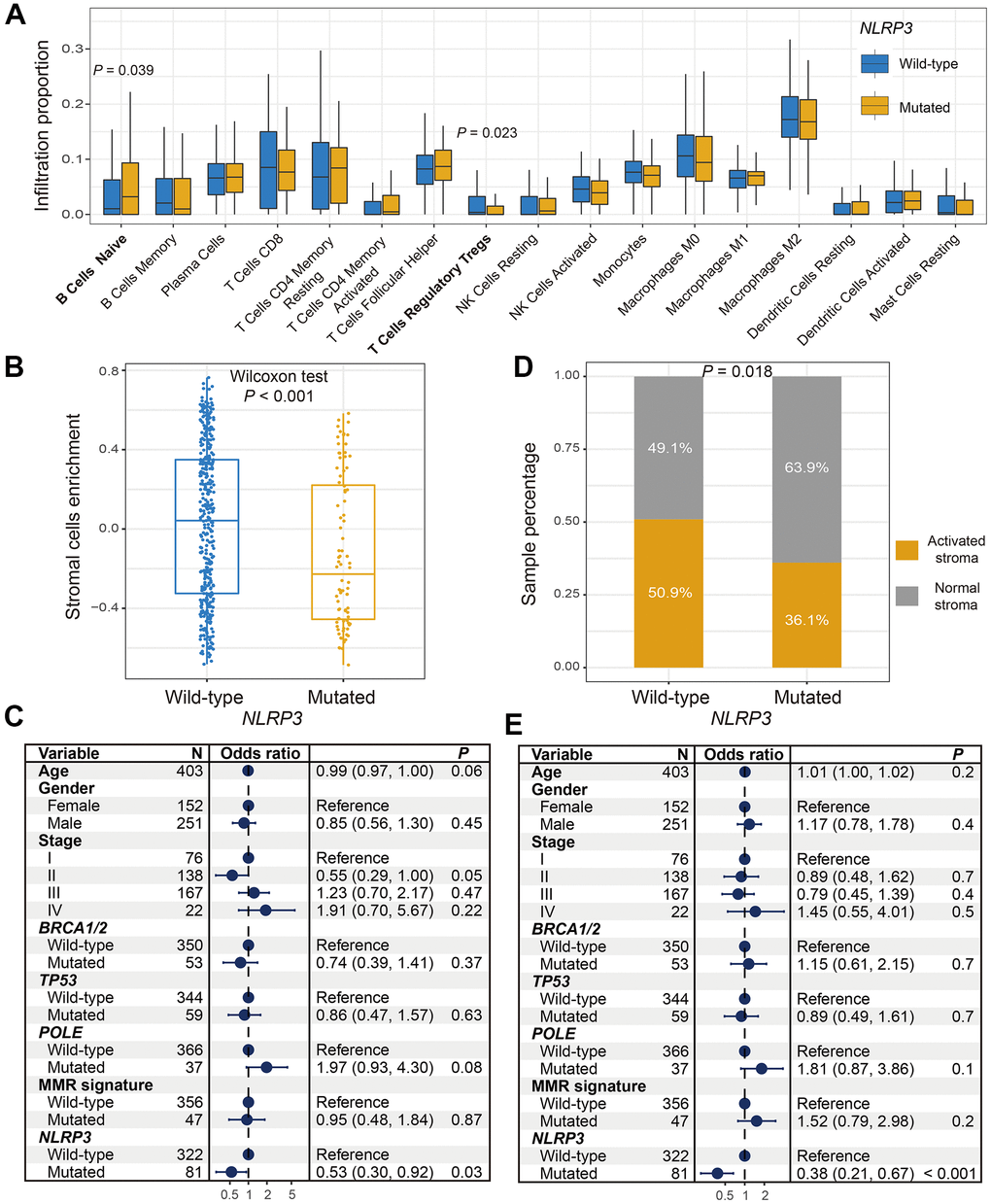
Figure 3. Association between NLRP3 mutations and factors in microenvironment. (A) Diverse infiltration abundance of immune cells based on NLRP3 mutational status. (B) Differential enrichment of overall stromal cells in NLRP3 mutated and wild-type patients. (C) Representation for forest plot of association between NLRP3 mutations and stromal cells enrichment. (D) Distinct distribution of activated stroma subtype in patients with and without NLRP3 mutations. (E) Multivariate Logistic regression model for the association of NLRP3 mutations with activated stroma subtype.
Stromal cells could promote tumor growth and inhibit immune response. Results indicated that patients with NLRP3 mutations harbored significantly lower enrichment of overall stromal cells (Wilcoxon rank-sum test, P < 0.001; Figure 3B). Multivariate Logistic regression model with other confounding factors taken into account still remained significant (OR: 0.53, 95% CI: 0.30-0.92, P = 0.03; Figure 3C). Proportion of activated stroma subtypes was significantly decreased in NLRP3 mutated patients as compared with wild-type patients (proportion: 36.1% vs. 50.9%, Fisher exact test, P = 0.018; Figure 3D). This result was more significant in multivariate analysis (OR: 0.38, 95% CI: 0.21-0.67, P < 0.001; Figure 3E).
Differential analyses of immune checkpoint genes showed that only CD276 exhibited a significantly elevated expression in NLRP3 mutated patients (Wilcoxon rank-sum test, P = 0.024; Supplementary Figure 4). Other checkpoint genes did not show statistical significance (Wilcoxon rank-sum test, all P > 0.05; Supplementary Figure 4). Collectively, the activated immune microenvironment was enriched in melanoma patients with NLRP3 mutations.
Clinical characteristics versus ICI efficacy in immunotherapy cohort
Before evaluating the association of NLRP3 mutations with ICI efficacy, we explored the influences of common clinical features (i.e., TMB, age, gender, stage, and treatment type) with 336 ICI-treated melanoma patients. We observed that high TMB was associated with elevated response rate (response rate: 35.7% vs. 25.0%, Fisher exact test, P = 0.043) and preferable overall survival (OS) (Log-rank test, P = 0.049) (Supplementary Figure 6A). Patients with age > 60 were likely to have a higher response rate than others (response rate: 41.4% vs. 23.7%, Fisher exact test, P = 0.002), but they did not exhibit a statistical difference in prognosis (Log-rank test, P = 0.942) (Supplementary Figure 6B). Male patients harbored a trend of high response rate, although it did not reach the statistical significance (response rate: 35.9% vs. 25.3%, Fisher exact test, P = 0.054); there is no significant difference in OS (Log-rank test, P = 0.286) (Supplementary Figure 6C). Patients with advanced-stage had the lowest response rate and worst prognosis, which may be correlated with their intrinsic properties (Supplementary Figure 6D). We found patients treated with anti-PD-1 therapy had the highest response rate than patients treated with anti-CTLA-4 or combined therapy (response rate: 41.6% vs. 31.8% vs. 16.3%, Fisher exact test, P = 0.002); nevertheless, patients who received combined therapy had the best OS (Log-rank test, P = 0.044) (Supplementary Figure 6E).
NLRP3 mutations were linked with favorable ICI efficacy
Associations of NLRP3 mutations with clinical characteristics among 336 ICI-treated melanoma patients were exhibited in Supplementary Table 1. Consistent with the aforementioned result, significantly increased TMB was identified in patients with NLRP3 mutations in the immunotherapy cohort (Wilcoxon rank-sum test, P < 0.001; Supplementary Figure 7A, 7B).
We found that NLRP3 mutated patients had significantly higher response rate than wild-type patients (response rate: 45.2% vs. 28.2%, Fisher exact test, P = 0.031; Figure 4A). Multivariate Logistic regression model also showed statistical difference after adjusting confounders (i.e., age, gender, stage, ICI therapy type, and TMB) (OR: 0.59, 95% CI: 0.28-1.25, P = 0.095; Figure 4B).
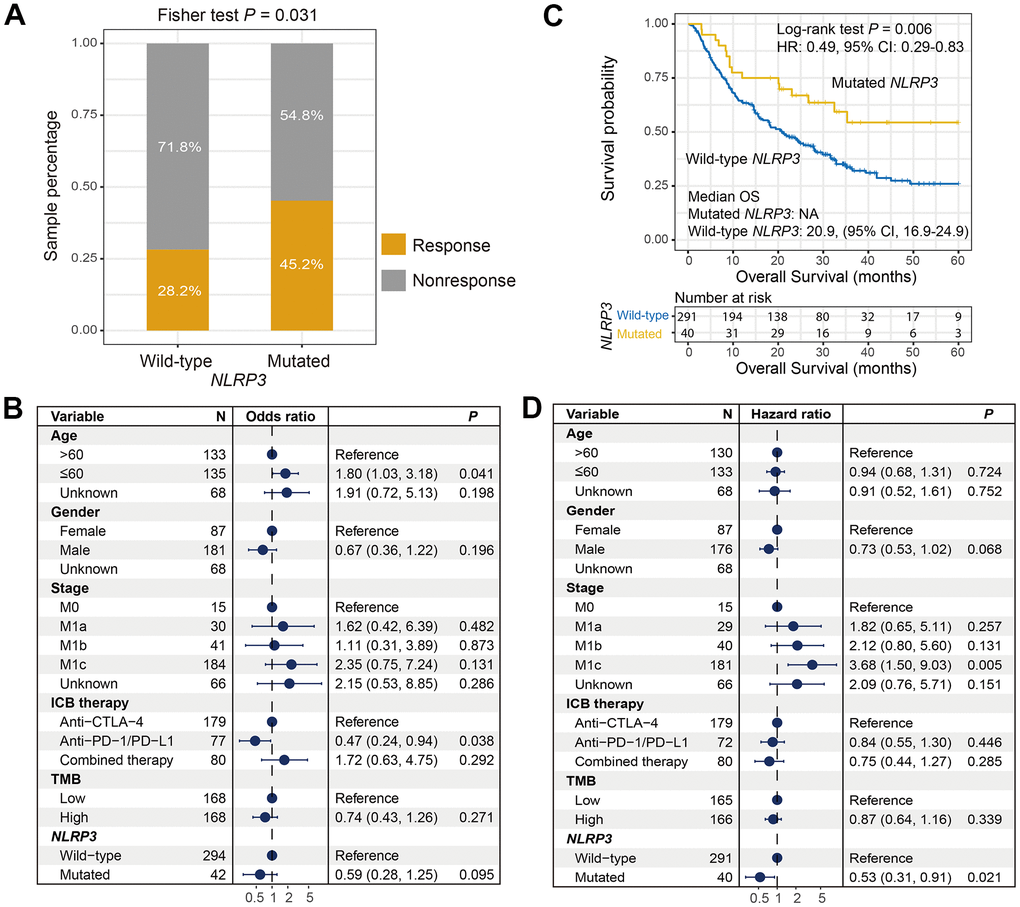
Figure 4. Correlation of NLRP3 mutations with ICI response rate and survival interval. (A) Distinct ICI response rate in NLRP3 mutated and wild-type patients. (B) Association of NLRP3 mutations with the response rate in multivariate Logistic regression model. (C) Kaplan-Meier survival curve of distinct NLRP3 status in ICI-treated cohort. (D) Forest plot for multivariate Cox regression model with confounders taken into account.
We calculated the intrinsic prognostic ability of NLRP3 mutations using 3 ICI-treated-naive cohorts from TCGA and ICGC. Results showed that NLRP3 mutations were not correlated with prognoses (Log-rank test, P = 0.461, P = 0.686 and P = 0.916; Supplementary Figure 8). However, survival analysis suggested that NLRP3 mutated patients had significantly preferable overall survival than wild-type patients in the ICI-treated cohort (median OS: not available [because more than half the patients in this subgroup were alive] vs. 20.9 [95% CI, 16.9-24.9], Log-rank test, P = 0.006; Figure 4C). Multivariate Cox regression analysis with clinical variables taken into account still remained significant (HR: 0.53, 95% CI: 0.31-0.91, P = 0.021; Figure 4D).
Finally, associations of NLRP3 mutations with ICI efficacy in distinct therapies were explored respectively. In patients treated with anti-CTLA-4 agents, no association was found between NLRP3 mutations and response rate (response rate: 38.1% vs. 31.1%, Fisher exact test, P = 0.685; Figure 5A); however, NLRP3 mutations were associated with better OS, although this result did not obtain statistical significance (Log-rank test, P = 0.063; Figure 5B). In patients treated with anti-PD-1 agents, we found that patients with NLRP3 mutations had a marginally significantly higher response rate (response rate: 70.0% vs. 36.4%, Fisher exact test, P = 0.081; Figure 5C); correlation of NLRP3 mutations with OS was not observed (Log-rank test, P = 0.568; Figure 5D). In patients received combined therapy, we also found NLRP3 mutant patients had a marginally significantly elevated response rate (response rate: 36.4% vs. 13.1%, Fisher exact test, P = 0.073; Figure 5E); and NLRP3 mutations were significantly correlated with prolonged immunotherapy OS (Log-rank test, P = 0.031; Figure 5F). Collectively, NLRP3 mutations were predictive of the preferable treatment efficacy in the settings of immunotherapy, especially the combined therapy.
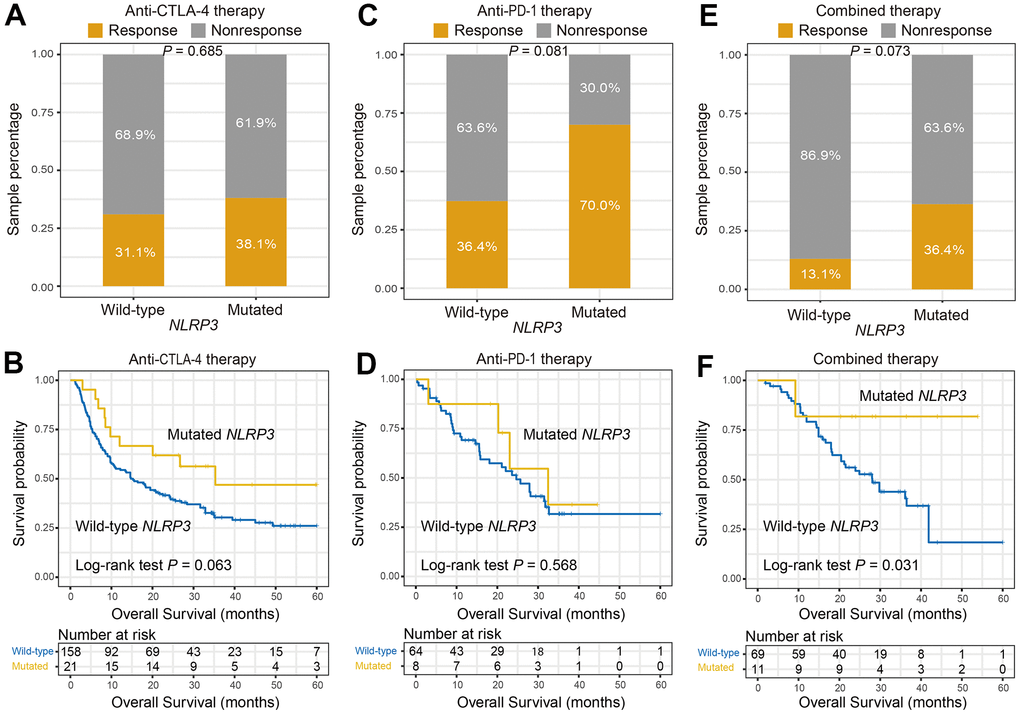
Figure 5. NLRP3 mutations association with ICI efficacy in distinct therapies. (A, B) Association of NLRP3 mutations with response rate and prognosis in patients treated with anti-CTLA-4 agents. (C, D) NLRP3 mutations versus response rate and prognosis in patients treated with anti-PD-1 agents. (E, F) NLRP3 mutations versus response rate and prognosis in patients who received combined therapy.
Discussion
By integrating and analyzing available genomic and clinical data of melanoma, NLRP3 mutations were identified to be associated with higher mutation and neoantigen burden, favorable microenvironment, and better tumor genomic features. Importantly, our study demonstrated that the elevated response and prolonged survival time of ICI therapy were found in patients with NLRP3 mutations. These findings suggest the predictive implications of NLRP3 mutations for melanoma immunotherapy.
Previously many studies revealed the vital roles of mutations of a single gene in evaluating ICI therapy efficacy. Jia et al. found that TTN mutations were positively associated with ICI predictive biomarkers and immunotherapy survival interval in melanoma and non-small cell lung cancer [26]. Patients with POLE/POLD1 mutations exhibited a significantly preferable prognosis in a multi-cancer-ICI cohort with 1644 patients [27]. In metastatic renal cell carcinoma patients who received Nivolumab antibody, Braun et al. observed that favorable overall and progression-free survival were markedly correlated with PBRM1 mutations [28]. High TMB and NB are 2 promising biomarkers in cancer immune treatment, however, some factors such as uncertain threshold, exome sequencing fees, and bias of different platforms largely influence the precise evaluation for both markers [26]. Mutations of NLRP3 could accurately assess high TMB and NB as our results described. Therefore, instead of choosing TMB and NB, NLRP3 mutations may be an alternative surrogate for predicting ICI response in melanoma.
Low tumor heterogeneity and purity suggest the reduced proportion of subclonal mutations and tumor cells in the microenvironment, which were reported to be correlated with better response to anti-PD-1 therapy [29]. Our study found that patients with NLRP3 mutations had decreased heterogeneity and purity, indicated the vital roles of NLRP3 mutations in immunotherapy response. The regulatory T cell is one immune cell subtype that performed immune-suppressive roles as well as stromal cells [30, 31]. NLRP3 mutations were found to be correlated with lower infiltration of regulatory T cells and stromal cells. Moreover, NLRP3 mutant patients harbored a reduced proportion of activated stroma subtype, which exhibits the roles of immune suppression. The above findings further verify the potentially positive association behind NLRP3 mutations and ICI therapy response.
In non-ICI-treated cohorts, no survival benefits were found in patients with NLRP3 mutations. Nevertheless, NLRP3 mutations exhibited a preferable response rate and ICI efficacy in the aggregated ICI cohort. Noticeably, for the roles of NLRP3 mutations in specific therapy, we found that NLRP3 mutated patients could obtain the best survival benefit during combined therapy as compared with anti-CTLA-4 and anti-PD-1 therapies. These findings suggest that NLRP3 mutations may serve as a predictive indicator for evaluating the efficacy of ICI, especially combined immunotherapy.
Recent research demonstrated that reduced NLRP3 inflammasome and IL-1β expression could inhibit the progression of cancer cells [24]. Consistently, another study reported that NLRP3 downregulation and reduced IL-1β secretion decreased metastatic melanoma by thymoquinone therapy in a mouse model [21]. The above findings suggested the crucial roles of NLRP3 and IL-1β in tumorigenesis and the development of melanoma. In this work, we also evaluated the markedly positive association of NLRP3 and IL-1β expression in melanoma (Supplementary Figure 9A) and further confirmed the collective roles of both regulators. Nevertheless, no significant differences were detected in NLRP3 wild-type and mutated subgroups with respect to NLRP3 and IL-1β expression (Supplementary Figure 9B, 9C). These results suggested that the mechanisms underlying the association between NLRP3 mutation and favorable immunotherapy efficacy may neither involve in NLRP3 nor IL-1β transcriptional signals.
A few limitations existed. Firstly, the gene expression-related analyses were performed with only the TCGA cohort, no additionally available datasets were used for validation. Secondly, biological relevance between NLRP3 mutations and immunological features was elusive, further studies were needed to explore.
In melanoma, NLRP3 mutations were associated with better immunological and genomic characteristics. It is worth noting that the elevated response rate and favorable ICI survival were also observed in NLRP3 mutated patients. NLRP3 mutations may harbor vitally predictive implications for immunotherapy response in melanoma.
Materials and Methods
Somatic mutation data, gene expression profile, and clinical information of included melanoma patients
Somatic mutation data of 467 melanoma patients in the Cancer Genome Atlas (TCGA) were derived from Genome Data Commons (https://portal.gdc.cancer.gov). MELA-AU and SKCA-BR cohorts respectively containing 183 and 100 patients derived from the International Cancer Genome Consortium (ICGC) (https://dcc.icgc.org) were utilized for specific validation. Gene expression data of 465 patients were acquired from the TCGA cohort.
From previously published 6 studies [1, 32–36], we collected a total of 336 pre-treatment advanced/metastatic melanoma patients with somatic mutation data. These patients were treated with anti-PD-1 agents, anti-CTLA-4 agents, or combined therapy in their trials. All somatic mutations were uniformly annotated with Oncotator [37]. Clinical information (e.g., age, gender, stage, and treatment type) and ICI efficacy (i.e., response status and survival time) of these 336 patients were described in Supplementary Table 2. In this study, patients with statuses of completed or partial response were defined as responders, other statuses including stable and progressive disease were not considered to be efficacious to ICI therapy.
Association of NLRP3 mutations with TMB, NB, and genomic features
Mutations of genomic maintenance genes were largely correlated with genome instability. Therefore, in addition to univariate analysis of the association of NLRP3 mutations with TMB and NB, we also conducted a multivariate Logistic regression model with mutations of DNA damage repair genes (i.e., TP53, BRCA1/2, and POLE) and mismatch repair (MMR) genes (i.e., MLH1, MSH2, MSH6, and PMS2) taken into account to control false positive. The neoantigen data of 340 patients were acquired from the Cancer Immunome Atlas (TCIA, https://www.tcia.at/home). TMB and NB were stratified into high and low subgroups with the median.
A number of studies have reported the vital roles of tumor genomic features (i.e., heterogeneity, purity, and ploidy) on immune response and immunotherapeutic efficacy [29, 38]. We therefore utilized relevant data of 140 patients derived from TCIA to evaluate the association of NLRP3 mutations with heterogeneity and ploidy. For the tumor purity of each patient, we used the ESTIMATE algorithm embedded in R package ESTIMATE [39] to calculate.
NLRP3 mutations versus tumor microenvironment
Overall infiltration of immune and stromal cells of each sample was evaluated with the aforementioned ESTIMATE algorithm. The nearest template prediction (NTP) algorithm [40] with a 48-gene signature [41] was applied to stratify melanoma patients into activated and normal stroma subgroups. A recent study reported that the activated stroma subtype exhibited an immune-suppressive role and a worse prognosis [41].
The infiltration abundance of 17 immune cell types was estimated with the CIBERSORT algorithm [42]. Angelova et al. established 812 immune metagene signatures to infer 31 immune cells infiltration and tumor immune landscape [43]. We used both methods to obtain comprehensive results and to validate each other.
An integrated list of 33 immune checkpoint genes was acquired from a recently published study [44]. We analyzed the distinct distributions of the above immune cells and checkpoint genes based on NLRP3 mutational statuses. All analyses in this section were performed with gene expression data of 465 samples from TCGA.
GSVA and GSEA
Single sample gene set enrichment analysis (ssGSEA) function embedded in GSVA package (V1.36.1) [45] was utilized to calculate the enrichment of a specific gene set for each patient. Differential analysis according to NLRP3 mutational statuses was performed with R package DESeq2 (V1.28.1) [46], which manages sequencing expression data. The t values obtained from differential analysis were subsequently used to conducted gene set enrichment analysis (GSEA) implemented by fgsea package (V1.14.0) (https://github.com/ctlab/fgsea). Kyoto encyclopedia of genes and genomes (KEGG) pathways were utilized as the background dataset.
Statistical analyses
R software (V4.0.1) and its affiliated packages were downloaded to complete related calculations and analyses. Waterfall plot of mutational patterns was achieved through GenVisR package (V1.20.0) [47]. Kaplan-Meier survival curve was drawn with survival (V2.41-3) and survminer (V0.4.7) packages and Log-rank test to compare the difference. We used forestmodel package (V0.5.0) to perform multivariate regression analyses and to produce forest plots. Correlations of NLRP3 mutations with continuous and categorical factors were calculated with Wilcoxon rank-sum test and Fisher exact test, separately.
Author Contributions
SW designed this study; SW, QW, and JL developed the methodology and acquired the related data; QW, JL, WZ, FS, YR, QM, YL, TL, and XL performed data analysis and interpretation; QW, SW, and JL drafted and revised the manuscript; SW supervised this study. All authors read and approved the final manuscript.
Acknowledgments
QHW would like to appreciate SZW at Weifang Medical University for her assistance in work and research. The authors thank Prof. Xiangchun Li at the Tianjin Medical University for the helpful suggestions.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This study was supported by the Shandong Provincial Youth Innovation Team Development Plan of Colleges and Universities (No. 2019-6-156, Lu-Jiao), National Natural Science Foundation of China (No. 81872719 and 81803337), Provincial Natural Science Foundation of Shandong Province (No. ZR201807090257), and National Bureau of Statistics Foundation Project (No. 2018LY79).
References
- 1. Roh W, Chen PL, Reuben A, Spencer CN, Prieto PA, Miller JP, Gopalakrishnan V, Wang F, Cooper ZA, Reddy SM, Gumbs C, Little L, Chang Q, et al. Integrated molecular analysis of tumor biopsies on sequential CTLA-4 and PD-1 blockade reveals markers of response and resistance. Sci Transl Med. 2017; 9:eaah3560. https://doi.org/10.1126/scitranslmed.aah3560 [PubMed]
- 2. Twitty CG, Huppert LA, Daud AI. Prognostic Biomarkers for Melanoma Immunotherapy. Curr Oncol Rep. 2020; 22:25. https://doi.org/10.1007/s11912-020-0886-z [PubMed]
- 3. Ribas A, Wolchok JD. Cancer immunotherapy using checkpoint blockade. Science. 2018; 359:1350–5. https://doi.org/10.1126/science.aar4060 [PubMed]
- 4. Robert C, Grob JJ, Stroyakovskiy D, Karaszewska B, Hauschild A, Levchenko E, Chiarion Sileni V, Schachter J, Garbe C, Bondarenko I, Gogas H, Mandalá M, Haanen JB, et al. Five-Year Outcomes with Dabrafenib plus Trametinib in Metastatic Melanoma. N Engl J Med. 2019; 381:626–36. https://doi.org/10.1056/NEJMoa1904059 [PubMed]
- 5. Larkin J, Chiarion-Sileni V, Gonzalez R, Grob JJ, Cowey CL, Lao CD, Schadendorf D, Dummer R, Smylie M, Rutkowski P, Ferrucci PF, Hill A, Wagstaff J, et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N Engl J Med. 2015; 373:23–34. https://doi.org/10.1056/NEJMoa1504030 [PubMed]
- 6. Havel JJ, Chowell D, Chan TA. The evolving landscape of biomarkers for checkpoint inhibitor immunotherapy. Nat Rev Cancer. 2019; 19:133–50. https://doi.org/10.1038/s41568-019-0116-x [PubMed]
- 7. Yarchoan M, Hopkins A, Jaffee EM. Tumor Mutational Burden and Response Rate to PD-1 Inhibition. N Engl J Med. 2017; 377:2500–1. https://doi.org/10.1056/NEJMc1713444 [PubMed]
- 8. Samstein RM, Lee CH, Shoushtari AN, Hellmann MD, Shen R, Janjigian YY, Barron DA, Zehir A, Jordan EJ, Omuro A, Kaley TJ, Kendall SM, Motzer RJ, et al. Tumor mutational load predicts survival after immunotherapy across multiple cancer types. Nat Genet. 2019; 51:202–6. https://doi.org/10.1038/s41588-018-0312-8 [PubMed]
- 9. Chan TA, Yarchoan M, Jaffee E, Swanton C, Quezada SA, Stenzinger A, Peters S. Development of tumor mutation burden as an immunotherapy biomarker: utility for the oncology clinic. Ann Oncol. 2019; 30:44–56. https://doi.org/10.1093/annonc/mdy495 [PubMed]
- 10. Sharma P, Callahan MK, Bono P, Kim J, Spiliopoulou P, Calvo E, Pillai RN, Ott PA, de Braud F, Morse M, Le DT, Jaeger D, Chan E, et al. Nivolumab monotherapy in recurrent metastatic urothelial carcinoma (CheckMate 032): a multicentre, open-label, two-stage, multi-arm, phase 1/2 trial. Lancet Oncol. 2016; 17:1590–8. https://doi.org/10.1016/S1470-2045(16)30496-X [PubMed]
- 11. Mangan MS, Olhava EJ, Roush WR, Seidel HM, Glick GD, Latz E. Targeting the NLRP3 inflammasome in inflammatory diseases. Nat Rev Drug Discov. 2018; 17:588–606. https://doi.org/10.1038/nrd.2018.97 [PubMed]
- 12. Moossavi M, Parsamanesh N, Bahrami A, Atkin SL, Sahebkar A. Role of the NLRP3 inflammasome in cancer. Mol Cancer. 2018; 17:158. https://doi.org/10.1186/s12943-018-0900-3 [PubMed]
- 13. Zaki MH, Boyd KL, Vogel P, Kastan MB, Lamkanfi M, Kanneganti TD. The NLRP3 inflammasome protects against loss of epithelial integrity and mortality during experimental colitis. Immunity. 2010; 32:379–91. https://doi.org/10.1016/j.immuni.2010.03.003 [PubMed]
- 14. Dupaul-Chicoine J, Yeretssian G, Doiron K, Bergstrom KS, McIntire CR, LeBlanc PM, Meunier C, Turbide C, Gros P, Beauchemin N, Vallance BA, Saleh M. Control of intestinal homeostasis, colitis, and colitis-associated colorectal cancer by the inflammatory caspases. Immunity. 2010; 32:367–78. https://doi.org/10.1016/j.immuni.2010.02.012 [PubMed]
- 15. Li S, Liang X, Ma L, Shen L, Li T, Zheng L, Sun A, Shang W, Chen C, Zhao W, Jia J. MiR-22 sustains NLRP3 expression and attenuates H. pylori-induced gastric carcinogenesis. Oncogene. 2018; 37:884–96. https://doi.org/10.1038/onc.2017.381 [PubMed]
- 16. Fan SH, Wang YY, Lu J, Zheng YL, Wu DM, Li MQ, Hu B, Zhang ZF, Cheng W, Shan Q. Luteoloside suppresses proliferation and metastasis of hepatocellular carcinoma cells by inhibition of NLRP3 inflammasome. PLoS One. 2014; 9:e89961. https://doi.org/10.1371/journal.pone.0089961 [PubMed]
- 17. Bae JY, Lee SW, Shin YH, Lee JH, Jahng JW, Park K. P2X7 receptor and NLRP3 inflammasome activation in head and neck cancer. Oncotarget. 2017; 8:48972–82. https://doi.org/10.18632/oncotarget.16903 [PubMed]
- 18. Sayan M, Mossman BT. The NLRP3 inflammasome in pathogenic particle and fibre-associated lung inflammation and diseases. Part Fibre Toxicol. 2016; 13:51. https://doi.org/10.1186/s12989-016-0162-4 [PubMed]
- 19. Veeranki S. Role of inflammasomes and their regulators in prostate cancer initiation, progression and metastasis. Cell Mol Biol Lett. 2013; 18:355–67. https://doi.org/10.2478/s11658-013-0095-y [PubMed]
- 20. Li L, Liu Y. Aging-related gene signature regulated by Nlrp3 predicts glioma progression. Am J Cancer Res. 2014; 5:442–9. [PubMed]
- 21. Ahmad I, Muneer KM, Tamimi IA, Chang ME, Ata MO, Yusuf N. Thymoquinone suppresses metastasis of melanoma cells by inhibition of NLRP3 inflammasome. Toxicol Appl Pharmacol. 2013; 270:70–6. https://doi.org/10.1016/j.taap.2013.03.027 [PubMed]
- 22. Guarda G, Zenger M, Yazdi AS, Schroder K, Ferrero I, Menu P, Tardivel A, Mattmann C, Tschopp J. Differential expression of NLRP3 among hematopoietic cells. J Immunol. 2011; 186:2529–34. https://doi.org/10.4049/jimmunol.1002720 [PubMed]
- 23. Drexler SK, Bonsignore L, Masin M, Tardivel A, Jackstadt R, Hermeking H, Schneider P, Gross O, Tschopp J, Yazdi AS. Tissue-specific opposing functions of the inflammasome adaptor ASC in the regulation of epithelial skin carcinogenesis. Proc Natl Acad Sci USA. 2012; 109:18384–9. https://doi.org/10.1073/pnas.1209171109 [PubMed]
- 24. Dunn JH, Ellis LZ, Fujita M. Inflammasomes as molecular mediators of inflammation and cancer: potential role in melanoma. Cancer Lett. 2012; 314:24–33. https://doi.org/10.1016/j.canlet.2011.10.001 [PubMed]
- 25. van Deventer HW, Burgents JE, Wu QP, Woodford RM, Brickey WJ, Allen IC, McElvania-Tekippe E, Serody JS, Ting JP. The inflammasome component NLRP3 impairs antitumor vaccine by enhancing the accumulation of tumor-associated myeloid-derived suppressor cells. Cancer Res. 2010; 70:10161–9. https://doi.org/10.1158/0008-5472.CAN-10-1921 [PubMed]
- 26. Jia Q, Wang J, He N, He J, Zhu B. Titin mutation associated with responsiveness to checkpoint blockades in solid tumors. JCI Insight. 2019; 4:e127901. https://doi.org/10.1172/jci.insight.127901 [PubMed]
- 27. Wang F, Zhao Q, Wang YN, Jin Y, He MM, Liu ZX, Xu RH. Evaluation of POLE and POLD1 Mutations as Biomarkers for Immunotherapy Outcomes Across Multiple Cancer Types. JAMA Oncol. 2019; 5:1504–6. https://doi.org/10.1001/jamaoncol.2019.2963 [PubMed]
- 28. Braun DA, Ishii Y, Walsh AM, Van Allen EM, Wu CJ, Shukla SA, Choueiri TK. Clinical Validation of PBRM1 Alterations as a Marker of Immune Checkpoint Inhibitor Response in Renal Cell Carcinoma. JAMA Oncol. 2019; 5:1631–3. https://doi.org/10.1001/jamaoncol.2019.3158 [PubMed]
- 29. Liu D, Schilling B, Liu D, Sucker A, Livingstone E, Jerby-Arnon L, Zimmer L, Gutzmer R, Satzger I, Loquai C, Grabbe S, Vokes N, Margolis CA, et al. Integrative molecular and clinical modeling of clinical outcomes to PD1 blockade in patients with metastatic melanoma. Nat Med. 2019; 25:1916–27. https://doi.org/10.1038/s41591-019-0654-5 [PubMed]
- 30. Ferreira LM, Muller YD, Bluestone JA, Tang Q. Next-generation regulatory T cell therapy. Nat Rev Drug Discov. 2019; 18:749–69. https://doi.org/10.1038/s41573-019-0041-4 [PubMed]
- 31. Denton AE, Roberts EW, Fearon DT. Stromal Cells in the Tumor Microenvironment. Adv Exp Med Biol. 2018; 1060:99–114. https://doi.org/10.1007/978-3-319-78127-3_6 [PubMed]
- 32. Van Allen EM, Miao D, Schilling B, Shukla SA, Blank C, Zimmer L, Sucker A, Hillen U, Foppen MH, Goldinger SM, Utikal J, Hassel JC, Weide B, et al. Genomic correlates of response to CTLA-4 blockade in metastatic melanoma. Science. 2015; 350:207–11. https://doi.org/10.1126/science.aad0095 [PubMed]
- 33. Hugo W, Zaretsky JM, Sun L, Song C, Moreno BH, Hu-Lieskovan S, Berent-Maoz B, Pang J, Chmielowski B, Cherry G, Seja E, Lomeli S, Kong X, et al. Genomic and Transcriptomic Features of Response to Anti-PD-1 Therapy in Metastatic Melanoma. Cell. 2016; 165:35–44. https://doi.org/10.1016/j.cell.2016.02.065 [PubMed]
- 34. Riaz N, Havel JJ, Makarov V, Desrichard A, Urba WJ, Sims JS, Hodi FS, Martín-Algarra S, Mandal R, Sharfman WH, Bhatia S, Hwu WJ, Gajewski TF, et al. Tumor and Microenvironment Evolution during Immunotherapy with Nivolumab. Cell. 2017; 171:934–49.e16. https://doi.org/10.1016/j.cell.2017.09.028 [PubMed]
- 35. Snyder A, Makarov V, Merghoub T, Yuan J, Zaretsky JM, Desrichard A, Walsh LA, Postow MA, Wong P, Ho TS, Hollmann TJ, Bruggeman C, Kannan K, et al. Genetic basis for clinical response to CTLA-4 blockade in melanoma. N Engl J Med. 2014; 371:2189–99. https://doi.org/10.1056/NEJMoa1406498 [PubMed]
- 36. Zaretsky JM, Garcia-Diaz A, Shin DS, Escuin-Ordinas H, Hugo W, Hu-Lieskovan S, Torrejon DY, Abril-Rodriguez G, Sandoval S, Barthly L, Saco J, Homet Moreno B, Mezzadra R, et al. Mutations Associated with Acquired Resistance to PD-1 Blockade in Melanoma. N Engl J Med. 2016; 375:819–29. https://doi.org/10.1056/NEJMoa1604958 [PubMed]
- 37. Ramos AH, Lichtenstein L, Gupta M, Lawrence MS, Pugh TJ, Saksena G, Meyerson M, Getz G. Oncotator: cancer variant annotation tool. Hum Mutat. 2015; 36:E2423–9. https://doi.org/10.1002/humu.22771 [PubMed]
- 38. Grzywa TM, Paskal W, Włodarski PK. Intratumor and Intertumor Heterogeneity in Melanoma. Transl Oncol. 2017; 10:956–75. https://doi.org/10.1016/j.tranon.2017.09.007 [PubMed]
- 39. Yoshihara K, Shahmoradgoli M, Martínez E, Vegesna R, Kim H, Torres-Garcia W, Treviño V, Shen H, Laird PW, Levine DA, Carter SL, Getz G, Stemke-Hale K, et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat Commun. 2013; 4:2612. https://doi.org/10.1038/ncomms3612 [PubMed]
- 40. Hoshida Y. Nearest template prediction: a single-sample-based flexible class prediction with confidence assessment. PLoS One. 2010; 5:e15543. https://doi.org/10.1371/journal.pone.0015543 [PubMed]
- 41. Moffitt RA, Marayati R, Flate EL, Volmar KE, Loeza SG, Hoadley KA, Rashid NU, Williams LA, Eaton SC, Chung AH, Smyla JK, Anderson JM, Kim HJ, et al. Virtual microdissection identifies distinct tumor- and stroma-specific subtypes of pancreatic ductal adenocarcinoma. Nat Genet. 2015; 47:1168–78. https://doi.org/10.1038/ng.3398 [PubMed]
- 42. Newman AM, Liu CL, Green MR, Gentles AJ, Feng W, Xu Y, Hoang CD, Diehn M, Alizadeh AA. Robust enumeration of cell subsets from tissue expression profiles. Nat Methods. 2015; 12:453–7. https://doi.org/10.1038/nmeth.3337 [PubMed]
- 43. Angelova M, Charoentong P, Hackl H, Fischer ML, Snajder R, Krogsdam AM, Waldner MJ, Bindea G, Mlecnik B, Galon J, Trajanoski Z. Characterization of the immunophenotypes and antigenomes of colorectal cancers reveals distinct tumor escape mechanisms and novel targets for immunotherapy. Genome Biol. 2015; 16:64. https://doi.org/10.1186/s13059-015-0620-6 [PubMed]
- 44. Ye Y, Jing Y, Li L, Mills GB, Diao L, Liu H, Han L. Sex-associated molecular differences for cancer immunotherapy. Nat Commun. 2020; 11:1779. https://doi.org/10.1038/s41467-020-15679-x [PubMed]
- 45. Hänzelmann S, Castelo R, Guinney J. GSVA: gene set variation analysis for microarray and RNA-seq data. BMC Bioinformatics. 2013; 14:7. https://doi.org/10.1186/1471-2105-14-7 [PubMed]
- 46. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014; 15:550. https://doi.org/10.1186/s13059-014-0550-8 [PubMed]
- 47. Skidmore ZL, Wagner AH, Lesurf R, Campbell KM, Kunisaki J, Griffith OL, Griffith M. GenVisR: Genomic Visualizations in R. Bioinformatics. 2016; 32:3012–14. https://doi.org/10.1093/bioinformatics/btw325 [PubMed]