




Introduction
Recently, nasopharyngeal carcinoma (NPC) has become a major health concern due to the high incidence in China, Alaska and North Africa [1]. Radiotherapy (RT) remains the first choice for the treatment of NPC, and the 5-year overall survival rate is about 80% [2]. For patients with locally advanced NPC, chemotherapy-radiotherapy (CRT) is undoubtedly the generally accepted treatment, with the capability of reducing the death rate by 18%, and enhancing the 5-year overall survival by 5% [3]. Due to the high metastatic characteristics of NPC, most patients have metastases by the time they are diagnosed [4]. The spread and metastasis of the NPC tumors results in a decreased survival rate of NPC patients and treatment failure, and currently there is no effective method to diagnose and hinder the development of NPC. Thus, a deeper understanding of the pathogenicity is critical to improve patient outcomes and reducing burden on healthcare systems.
Recently, microRNAs (miRNAs) have been reported to be involved in the regulatory mechanisms of many diseases, particularly in human cancers [5]. The roles of miR-212 in different cancers have drawn more and more attention. Downregulation of miR-212 has been identified in colorectal carcinoma [6], gastric cancer [7], and lung carcinoma [8]. Functionally, miR-212 was found to inhibit the progression of multiple cancers, including osteosarcoma [9], gastric cancer [10] and ovarian cancer [11]. However, carcinogenic effect of miR-212 was also detected in other cancers. For example, enhanced expression of miR-212 was detected in pancreatic ductal adenocarcinoma, and it was determined that increased expression promoted cell invasion and tumor growth [12]. These findings imply that distinct roles of miR-212 depend on the types of cancer. The role of miR-212 in the progression of NPC has not yet been fully explored.
The epithelial–mesenchymal transition (EMT) is recognized as a transition in which epithelial cells acquire the features of mesenchymal stem cells [13]. A critical feature of EMT is the decreased expression of epithelial cadherin (E-cadherin), coupled with the de novo or overexpression of neural cadherin (N-cadherin) [14]. Vimentin is another EMT marker protein. It is overexpressed in various epithelial cancers, and correlates well with enhanced tumor proliferation, invasion, and poor prognosis [15]. Moreover, the Wingless and Int-1 (Wnt)/β-catenin pathways could regulate many vital cellular processes such as stem cell regeneration and organogenesis [16]. Wnt signaling activation has been detected in lung, hematopoietic, and lung malignancies and is related to the recurrence of tumor. The Wnt/β-catenin pathway and the epithelial–mesenchymal transition (EMT) are both involved in tumorigenesis of NPC [13, 16]. miR-212 has previously been demonstrated to regulate EMT and the Wnt/β-catenin pathway in prostate cancer and hepatocellular carcinoma [17, 18]. Whether miR-212 is involved in NPC has been unclear.
The dysregulation of tumor protein D52 (TPD52) is a short hydrophilic polypeptide of 19.8 kDa. This protein is proposed to participate in cell proliferation and tissue invasion, and has been associated with various cancers. The enhanced expression of TPD52 has been recognized to be an early event in breast, prostate and ovarian cancer [19–21]. It was also suggested that increased TPD52 expression may be a marker of tumor susceptibility or very early precancerous lesions [22]. High expression of TPD52 may also promote the occurrence of primary cancer [21]. The high expression of TPD52 in the early and late stages of cancer suggests that it may be involved in tumor initiation and progression through independent mechanisms. Upregulation of TPD52 has been closely correlated to histological subtypes and patient prognosis in ovarian cancer [21]. In addition, downregulation of TPD52 induced by the anti-tumor miRNA miR-218 has been found to suppress the progression of lung squamous cell carcinoma [23]. Although the carcinogenic effects of TPD52 have been extensively investigated, its role in NPC has not been reported.
In this study, the alternation of miR-212 expression in NPC was examined, the correlation of miR-212 level and clinical features of NPC patients was analyzed. We found miR-212 reduces the expression of TPD52 through targeting its 3’ UTR, and thus inhibits EMT and Wnt/β-catenin pathway in NPC. This study expands our knowledge of the role of microRNAs in nasopharyngeal carcinoma and provides a promising novel treatment option for NPC.
Results
Downregulation of miR-212 in nasopharyngeal carcinoma patient tissues and cell lines
The level of miR-212 was explored in NPC cell lines and patient tissues by qRT-PCR. The relative level of miR-212 normalized to U6 in NPC tissues was decreased compared with normal tissues (p<0.01, Figure 1A). Similarly, the level of miR-212 was also observed lower in C666-1 and 6-10B NPC cell lines relative to the normal NP69 cells (p<0.01, Figure 1B). Moreover, the tumor/node/metastasis (TNM) stage (p = 0.017) and distant metastasis (p = 0.022, Table 1) in NPC patients were associated with downregulation of miR-212, and NPC patients expressing lower levels of miR-212 had a decreased overall survival (p = 0.0436, Figure 1C). We also analyzed miR-212 expression in NPC and normal tissues in public available study cohorts in TCGA database and found low miR-212 in NPC tissues (p = 0.0036, Figure 1D). These results thus indicated that NPC is associated with lower level of miR-212.
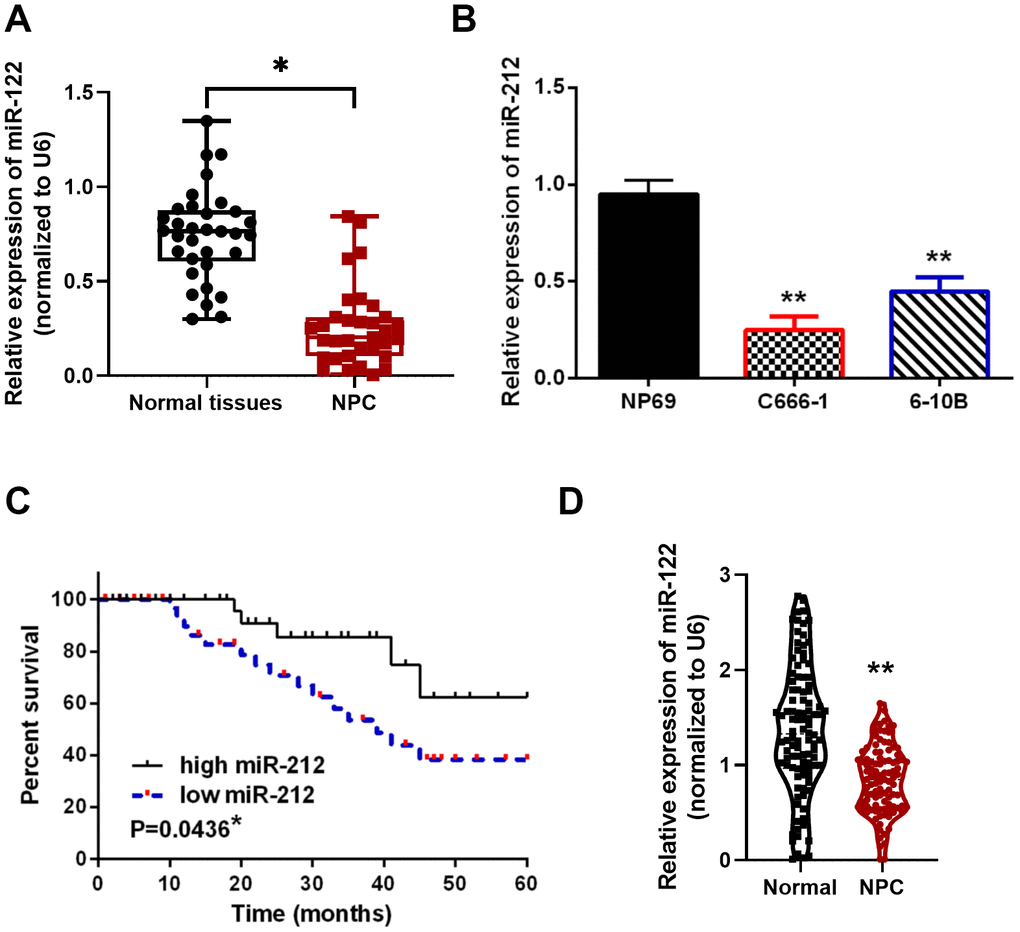
Figure 1. Downregulation of miR-212 in NPC tissues and cell lines. (A) Low miR-212 expression was observed in NPC tissues. N=33 in each group. (B) Relative MiR-212 expression levels in C666-1, 6-10B and NP69 cell lines. N=4 in each group. (C) Low miR-212 expression was associated with worse NPC prognosis. (D) Low miR-212 expression in NPC was found in public available study cohorts in TCGA database (N=102). *p < 0.05, ** p <0.01.
Table 1. The relationship between miR-212 expression and clinic-pathological characteristics of NPC patients.
| Characteristics | Cases(n=58) | miR-212 | P-value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| High(n=19) | Low(n=39) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (years) | 0.24 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥ 50 | 34 | 12 | 22 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <50 | 24 | 7 | 17 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gender | 0.69 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Male | 32 | 9 | 23 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Female | 26 | 10 | 16 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TNM stage | 0.017* | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| I+II | 19 | 6 | 13 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| III+IV | 39 | 13 | 26 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Distant metastasis | 0.022* | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| No | 42 | 14 | 28 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Yes | 16 | 5 | 11 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lymph node metastasis | 0.73 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| No | 38 | 11 | 27 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Yes | 20 | 8 | 12 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Statistical analyses were performed by the χ2 test. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *P<0.05 was considered significant. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Upregulation of miR-212 in NPC cells hindered cell migration, proliferation and invasion
To pinpoint the effects of miR-212 on NPC pathogenesis, miR-212 mimics or inhibitors were transfected into C666-1 cells. MiR-212 expression was observed to be regulated by miR-212 mimics or inhibitor, indicating that transfection was effective (p<0.01, Figure 2A). Functionally, miR-212 overexpression inhibited proliferation of C666-1 cells. In contrast, miR-212 downregulation promoted cell proliferation in NPC cells (p<0.05 or 0.01, Figure 2B). Moreover, miR-212 mimics inhibited migration and invasion of C666-1 cell while the miR-212 inhibitor promoted cell migration and invasion as shown by transwell assay (p<0.01, Figure 2C, 2D). Similarly, miR-212 overexpression inhibited cell migration whereas downregulation of miR-212 promoted C666-1 cell migration in the wound healing assay (p<0.01, Figure 2E). Thus, these data suggested that miR-212 possesses an inhibitory capacity in the context of NPC tumorigenesis.
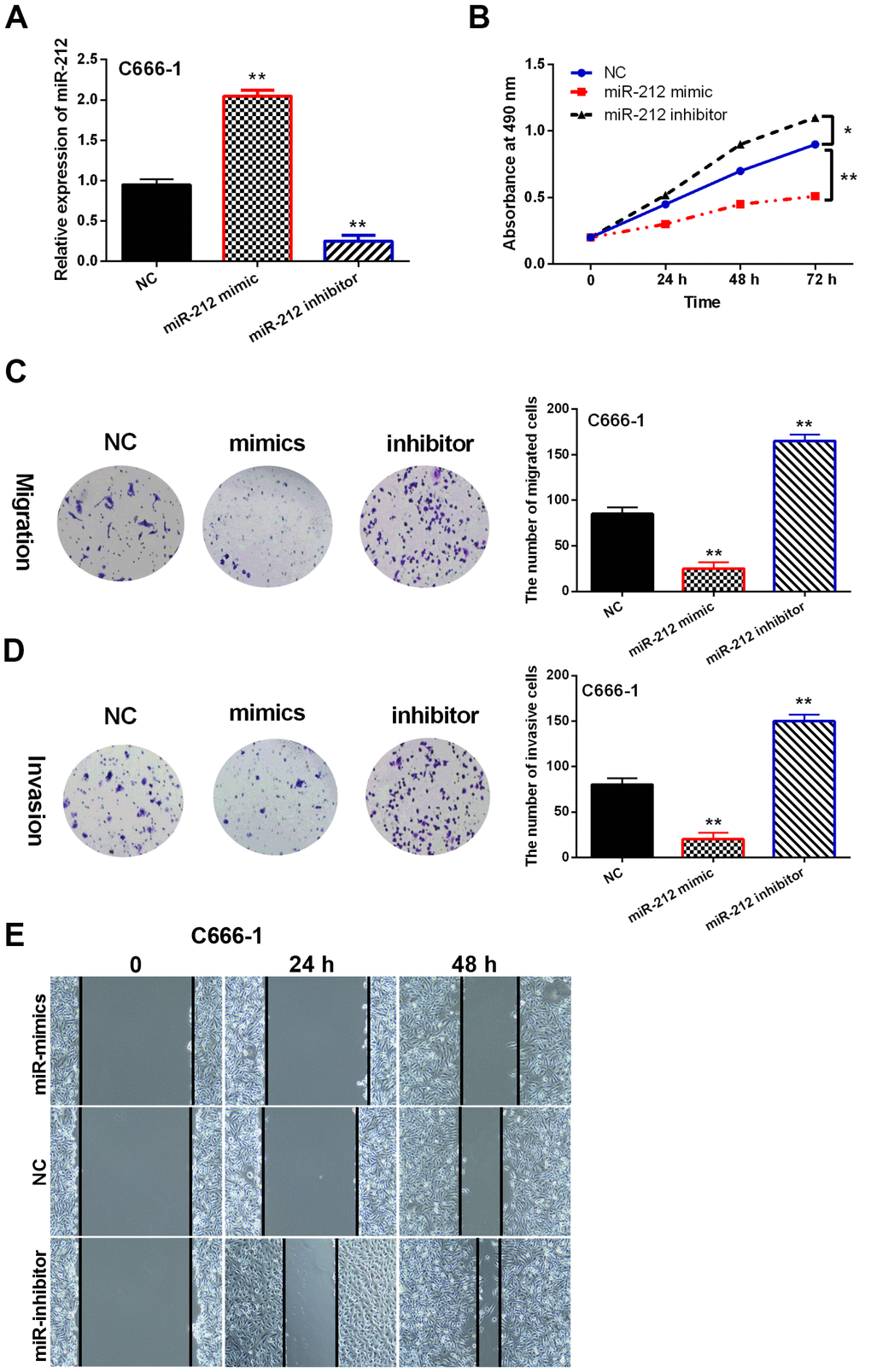
Figure 2. MiR-212 overexpression inhibited migration and proliferation of NPC. NC represented C666-1 cells with the negative control sequence. (A) Effect of miR-212 mimics or inhibitor on miR-212 expression in C666-1 cells. N=3. (B) miR-212 mimics or inhibitor influenced cell proliferation in C666-1 cells as detected by MTT assay. N=3. (C, D) miR-212 mimics or inhibitor regulated the migration and invasion of C666-1 cells as detected by Transwell assay. Representative images were shown (Magnification, 40×). Bar graphs indicate mean±SD of 3 independent experiments. (E) Wound healing assay of C666-1 cells transfected with NC, miR-mimics and miR-inhibitor. *p <0.05, **p <0.01.
MiR-212 inhibited EMT and inactivated the Wnt/β-catenin pathway in NPC cells
To gain a deeper understanding of the regulatory functions of miR-212, we sought to determine how miR-212 regulates EMT and the Wnt/β-catenin pathway in NPC. Overexpression of miR-212 in C666-1 and 6-10B cells reduced protein levels of EMT indicators, including Vimentin and N-cadherin but increased the amount of E-cadherin protein (Figure 3A). Unsurprisingly, knockdown of miR-212 led to increased Vimentin and N-cadherin expression while reducing the expression of E-cadherin (Figure 3A).
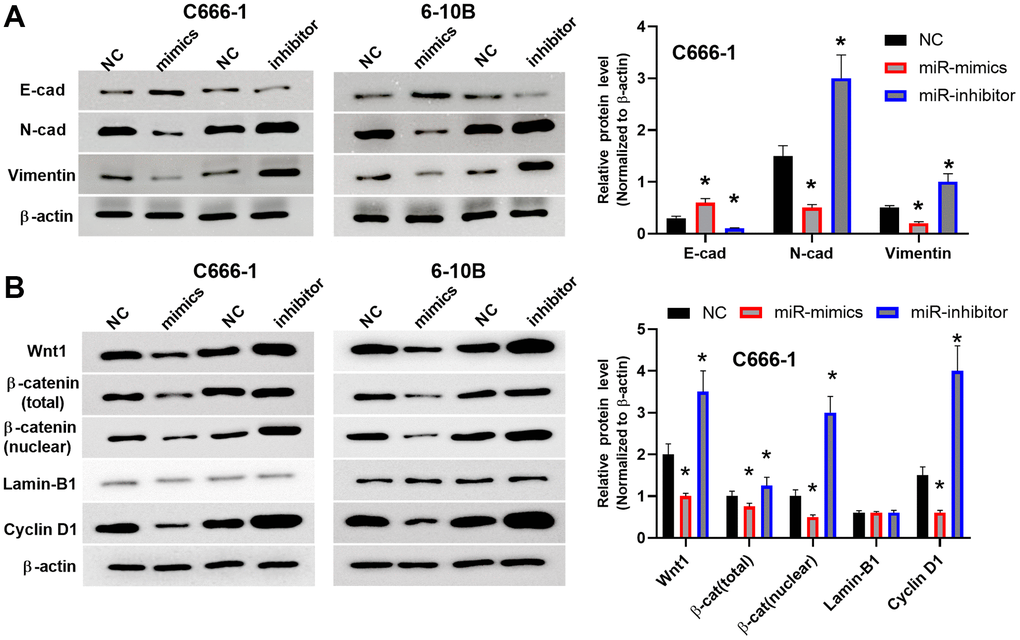
Figure 3. MiR-212 inhibited EMT and inactivated the Wnt/β-catenin pathway in NPC cells. (A) The protein levels of Vimentin, N-cadherin and E-cadherin were regulated by miR-212 mimics or inhibitor in C666-1 cells. NC means C666-1 cells with empty sequence. n=3-4. Data were quantified and normalized to the intensity of β-actin (left). (B) The protein levels of Wnt/β-catenin and downstream Cyclin D1 were analyzed by western blot. Both total β-catenin and nuclear localized β-catenin were shown. Lamin-B1 was used as the loading control for nuclear proteins. The western blots (right) were the representative images of 3-4 experiments. Data were quantified and normalized to the intensity of β-actin (left). *p <0.05, **p <0.01.
Because the Wnt/β-catenin pathway is engaged in the pathogenesis of cancers, we proposed that the Wnt/β-catenin pathway may also influence NPC progression. In Figure 3B, the expression of Wnt/β-catenin and downstream Cyclin D1 were studied by western blot. We showed both total β-catenin and nuclear localized β-catenin because the translocation of β-catenin from the cell membrane or the cytoplasm to the nuclear is an indicator of the canonical Wnt signaling pathway activation. We also analyzed the protein level of Cyclin D1 since it is a downstream target gene of the Wnt/β-catenin signaling pathway. High levels of Cyclin D1 protein promotes cell proliferation. Our data indicated downregulation of miR-212 activated Wnt/β-catenin pathway which may promote the proliferation of NPC. We found overexpression of miR-212 inhibited the expression of Wnt1, total β-catenin, nuclear β-catenin and the downstream protein Cyclin D1; while downregulation of miR-212 promoted the expression of Wnt1, β-catenin and Cyclin D1. Overall, these data indicated that miR-212 suppress the progression of NPC by impeding both EMT and the Wnt/β-catenin pathway.
MiR-212 targeting TPD52
To further investigate the mechanism by which miR-212 inhibits the formation of NPC, we searched for additional targets of miR-212 using the TargetScan database. It was predicted that TPD52 containing the binding sites of miR-212 (Figure 4A). To verify this prediction, we designed a luciferase reporter assay to test the interaction of miR-212 in the 3’ UTR of TPD52. Overexpression of miR-212 reduced WT-TPD52 luciferase activity, but had no impact on Mut-TPD52 luciferase activity (p<0.01, Figure 4B). Furthermore, miR-212 expression was negatively associated with TPD52 expression in NPC tissues (p<0.0001, R2=0.7522; Figure 4C). To further verify this antagonistic relationship, the impact of miR-212 on TPD52 expression was measured in C666-1 and 6-10B cells following overexpression or knockdown of miR-212 (Figure 4D). We found that TPD52 was downregulated by miR-212 mimics and upregulated by miR-212 inhibitor (p<0.01). Overall, these data demonstrated that miR-212 directly bound to TPD52 mRNA and subsequently caused a decrease in TPD52 expression in NPC.
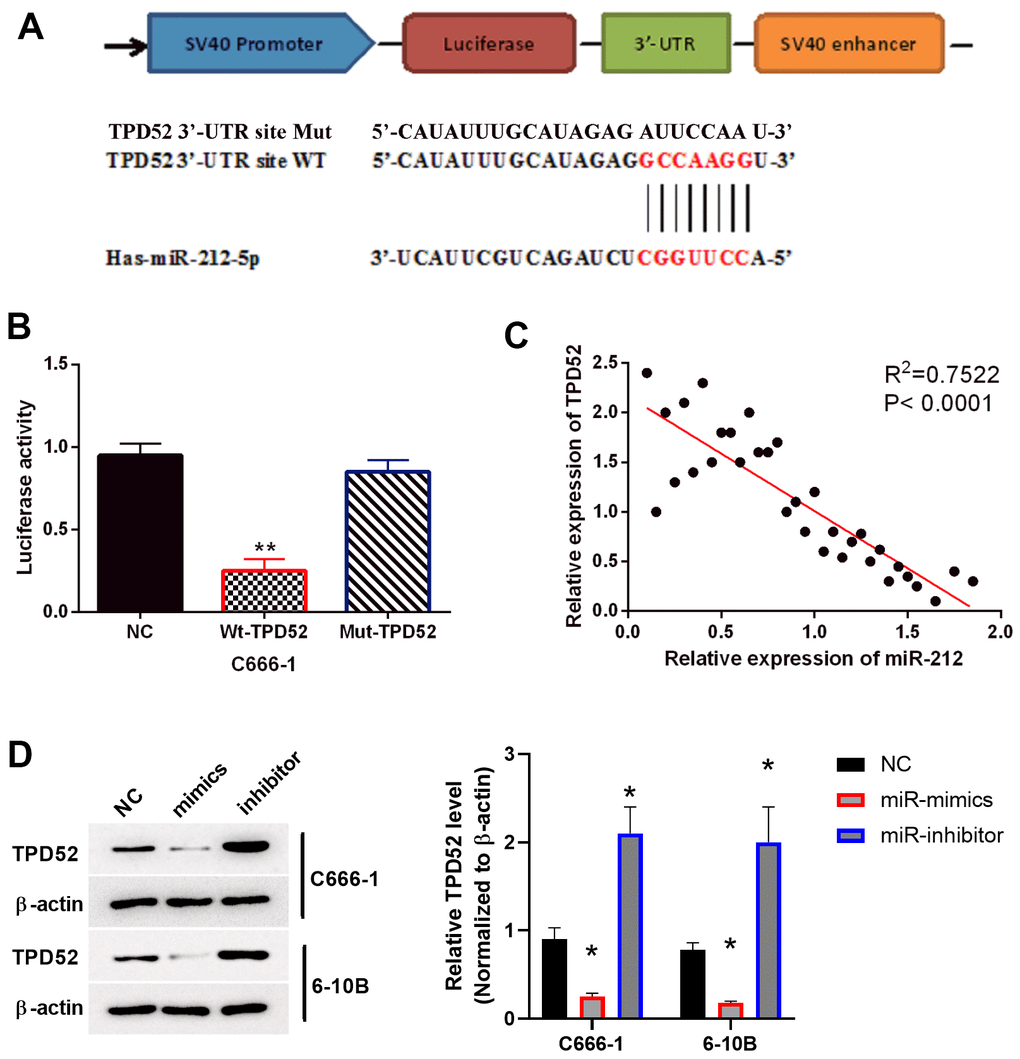
Figure 4. TPD52 was a direct target of miR-212. NC represented C666-1 cells with empty sequence. (A) Predicted binding site between TPD52 and miR-212. (B) Luciferase reporter assay with WT-TPD52 and Mut-TPD52. N=3. (C) miR-212 was negatively correlated with TPD52 in NPC tissues. N=33. (D) miR-212 mimics or inhibitor regulated TPD52 expression. The western blots were the representative images of 3 experiments. Data were quantified and normalized to the intensity of β-actin. **p <0.01.
MiR-212 inhibited NPC progression via targeting TPD52
Through qPCR, TPD52 expression was found elevated in NPC tissues (p<0.01, Figure 5A). The protein levels of TPD52 were also increased in NPC tissues from patients (p<0.01, Figure 5B). Moreover, upregulation of TPD52 correlated with the poor prognosis in NPC patients (p=0.0357, Figure 5C). High TPD52 expression in NPC was found in publicly available study cohorts in TCGA database (p<0.01, Figure 5D). We also detected the enhanced expression of TPD52 in C666-1 and 6-10B cell lines (p<0.05, Figure 5E). These results indicate that TPD52 may be involved in NPC progression. Next, miR-212 mimics suppressed the expression of TPD52, while miR-212 inhibitor enhanced the expression of TPD52 in C666-1 cells, further confirmed their interaction in NPC (p<0.05, Figure 5F).
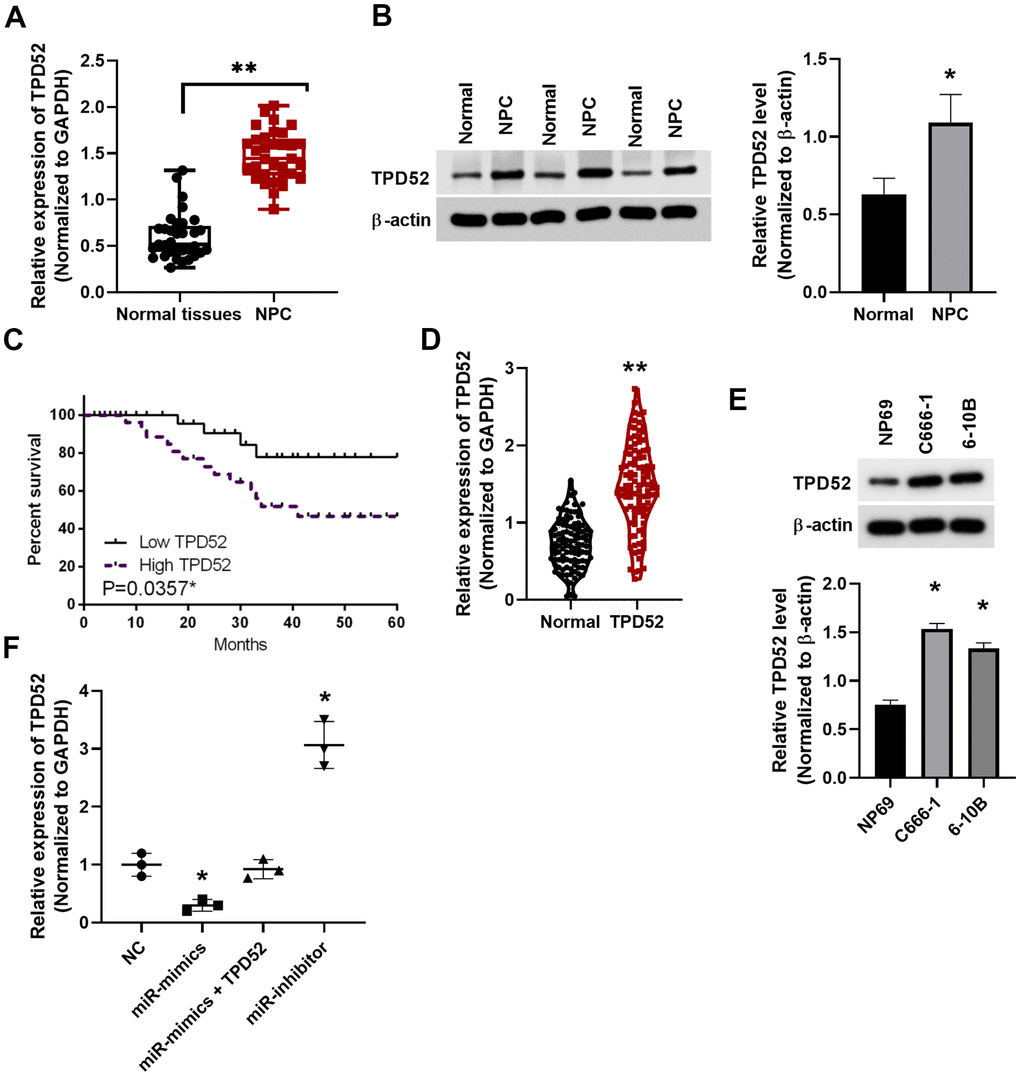
Figure 5. Upregulation of TPD52 in NPC patient tissue and cell lines. (A) TPD52 expression was analyzed in NPC tissues via qRT-PCR. N=33. (B) TPD52 protein levels were analyzed by western blot and normalized to β-actin (N=10). (C) Low TPD52 expression predicted good prognosis in NPC patients. (D) High TPD52 expression in NPC was found in public available study cohorts in TCGA database (N=102). (E) High TPD52 protein levels in NPC cell lines (N=3). (F) TPD52 expression was measured in C666-1 cells transfected with miR-212 mimics and inhibitor (N=3). *p <0.05, **p <0.01.
Functionally, the previously observed inhibition of cell proliferation induced by miR-212 mimics was impaired by overexpression of TPD52 (Figure 6A). Similar results were observed for the transwell assay of cell migration (Figure 6B) and invasion (Figure 6C), as well as the wound healing assay in C666-1 cells (Figure 6D). siRNA knockdown of TPD52 inhibited EMT and Wnt/β-catenin pathway, as shown by western blot of marker proteins in C666-1 cells (Figure 6E, 6F). On the contrary, overexpression of TPD52 stimulated the expression of EMT and Wnt/β-catenin proteins. Overall, these results suggested that miR-212 suppressed cell viability and EMT in NPC cells through targeting TPD52.
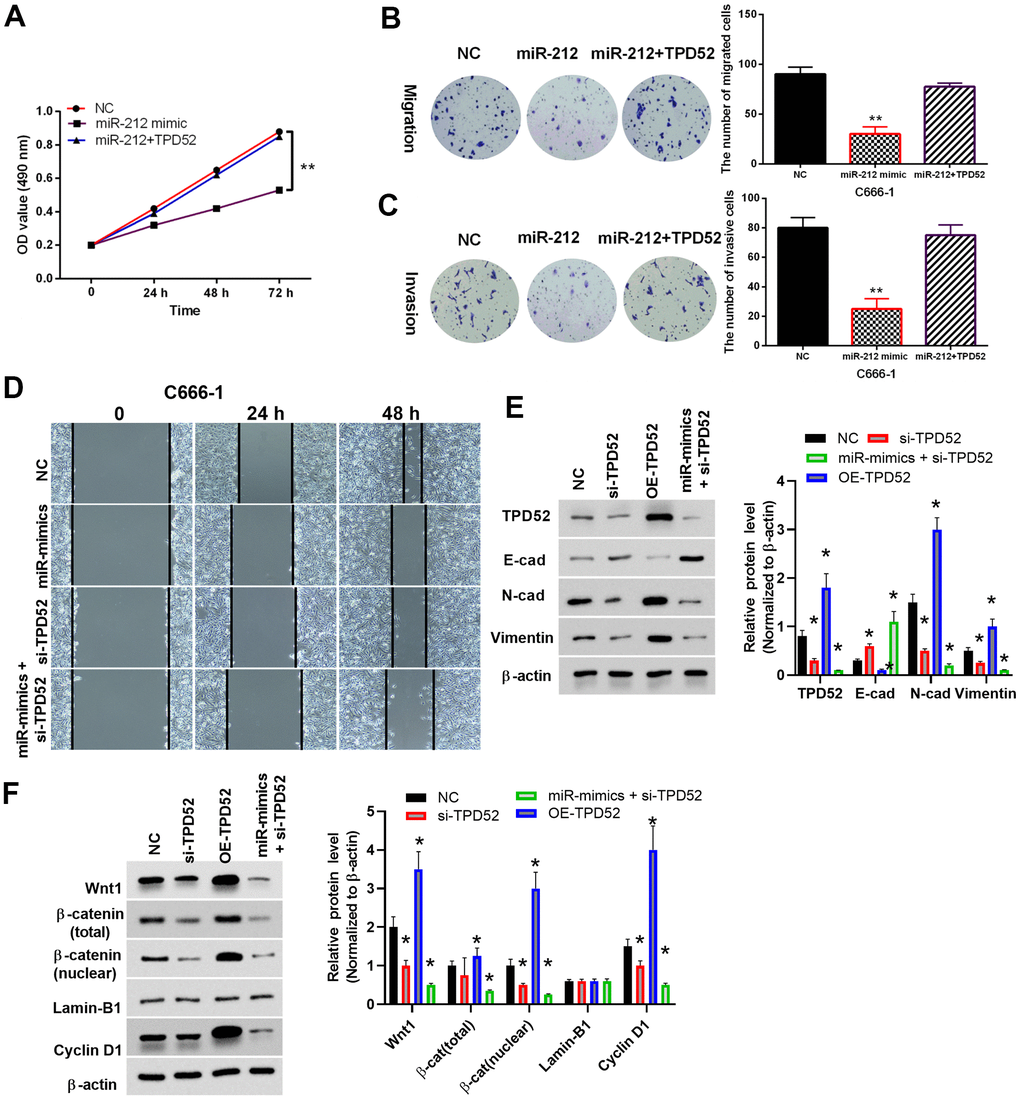
Figure 6. MiR-212 inhibited NPC through targeting TPD52. NC represented cells with empty sequence. (A) The effect of TPD52 on cell proliferation. N=3. (B, C) The effect of TPD52 on cell migration and invasion as shown by Transwell assay. Representative images were shown (Magnification, 40×). Bar graphs indicate mean±SD of 3 independent experiments. (D) The effect of TPD52 on cell migration as shown by wound healing assay. (E, F) Representative western blot analysis and quantified bar-graphs of EMT marker proteins and Wnt/β-catenin pathway proteins under up- or downregulation of TPD52. *p <0.05, **p <0.01.
MiR-212 inhibited tumor growth in vivo
In order to validate the effects of miR-212 on TPD52 expression and NPC tumor growth in vivo, the transplanted tumor model with miR-212 up- and downregulation were established in nude mice. Tumor volumes were compared among different groups. We found that the volume of C666-1 tumors with overexpression of miR-212 were considerably lower than those of the control (Figure 7A), and tumor volumes with miR-212 inhibition were much higher than the control, indicating miR-212 overexpression inhibited tumor growth. In order to confirm whether miR-212 regulates the TPD52 and Wnt/β-catenin pathway in vivo, western blot was performed using tumor tissues from different groups (Figure 7B). The result showed that TPD52 and Wnt/β-catenin pathway were downregulated under miR-212 overexpression in tumors in vivo. Immunohistochemical staining of tumor tissues indicated inhibition of miR-212 enhanced proliferation and invasion of tumor cells, as revealed by Ki-67 and CD31 staining, respectively (Figure 7C). The results suggested that miR-212 downregulated TPD52 and Wnt/β-catenin pathway and inhibited tumor growth in vivo.
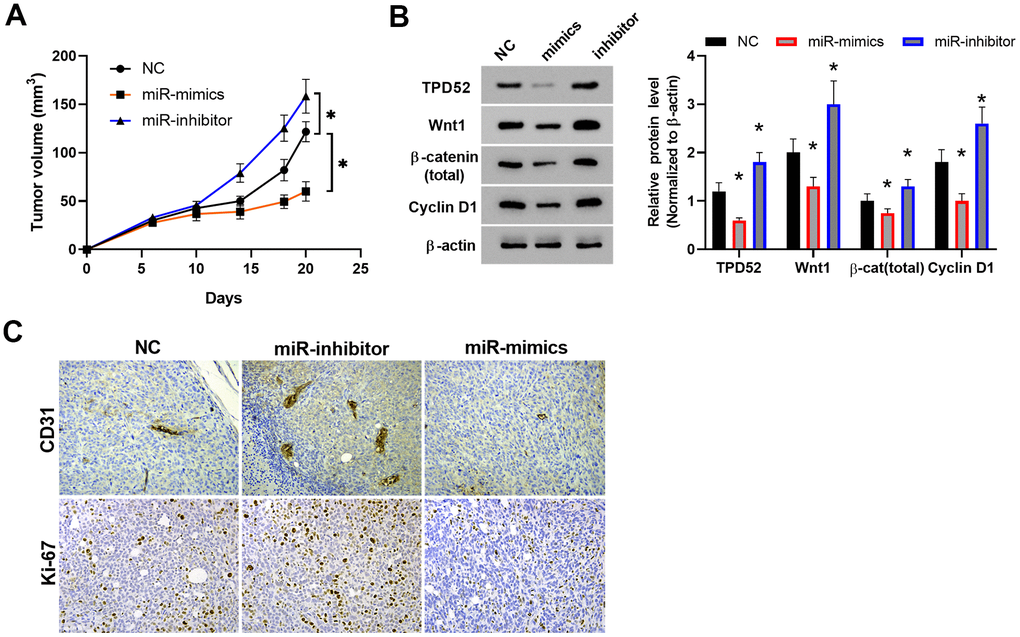
Figure 7. MiR-212 inhibited tumor growth in vivo. (A) Tumor volume growth curve. n=6. (B) The expression of TPD52 and Wnt/β-catenin were detected by western blot. n=6. (C) The proliferation and angiogenesis of tumor tissue section as detected by Ki-67 and CD31 immunostaining, respectively. Representative images were shown. * p < 0.05; ** p < 0.01.
Discussion
More and more miRNAs have been detected in the development of NPC, and studies showed that abnormal expressions of miRNAs can promote or inhibit the occurrence of NPC tumors. These findings reveal the roles of potential miRNAs in the diagnosis and treatment of NPC. For example, downregulation of miR-324 was detected in NPC and was found to be associated with clinical significance [24]. In this study, we revealed that miR-212 was downregulated in NPC, and the decreased expression was associated with important clinical developments such as distant metastasis and TNM stage in NPC patients. We also found that NPC cell migration, proliferation and invasion were inhibited by upregulation of miR-212. Our results indicated that miR-212 may inhibit metastasis by blocking EMT in NPC cells. These findings revealed that miR-212 is an inhibitory miRNA in the development of NPC.
Consistent with our results, decreased miR-212 expression was found in gastric cancer [25], and it has been demonstrated that miR-212 can suppress cell viability and metastasis in renal cell carcinoma [26]. Furthermore, miR-212 was found to suppress EMT in breast cancer [27]. Furthermore, miR-212 has been found to be involved in tumorigenesis through targeting several genes, such as Sox4 and SGK3 [28, 29]. In the current study, we are the first to find that miR-212 acts through a similar mechanism by directly targeting TPD52 mRNA for degradation.
miR-212 levels were found to be negatively correlated with TPD52 expression in NPC tissues. Overexpression of TPD52 impaired the inhibitory effect of miR-212 in NPC. TPD52 was recognized to be a target gene regulated by several miRNAs, such as miR-224 and miR-139 [30, 31], and upregulation of TPD52 has been identified in breast cancer [32]. In this study, increased TPD52 expression was observed in NPC and demonstrated a positive correlation with poor patient prognosis. Previous studies have investigated additional interactions between TPD52 and other miRNAs in various human cancers. For example, miR-34a inhibited cell metastasis by suppressing oncogenic TPD52 in breast cancer [33], and miR-379 inhibited cell viability and metastasis via targeting TPD52 in NPC [34]. Similarly, miR-212 also inhibited the progression of NPC through targeting TPD52.
TPD52 is regulated by multiple miRNAs such as miR-34a and miR-499b-5p [33, 35]. These miRNAs are further regulated by LncRNAs such as LncRNA NEAT1, which targeting miR-34a-5p and driving NPC progression [36]. Thus, there was a complicated interaction network of multiple miRNAs and TPD52 in the progression of cancer. The context and microenvironment of tumor cells may lead to the activation or inhibition of certain regulation pathways in vivo. Besides miR-212, our preliminary study showed that both miR-34a and miR-499b-5p were slightly downregulated (about 1.4-1.8 fold) in C666-1 cell compared to the NP69 cells. It future studies, it would be interesting to characterize which one is critical for the regulation of TPD52 protein level in NPC.
Furthermore, we reported that miR-212 significantly affects the Wnt/β-catenin pathway. Previous reports have shown that enhanced level of β-catenin is related to progression of malignant tumor [37]. Our study demonstrated that miR-212 overexpression reduced the level of β-catenin, indicating that miR-212 acts to inactivate the Wnt/β-catenin pathway in NPC. Consistent with our findings, upregulation of miR-212 has been reported to deactivate the Wnt/β-catenin pathway in hepatocellular carcinoma [18]. These studies indicate the potential of miR-212 for tumor suppression in NPC.
Conclusions
This study proved first-time that miR-212 was lower in NPC and that decreased level of miR-212 was involved in the malignant of NPC. Furthermore, miR-212 exerted an inhibitory effect on NPC progression through targeting TPD52 and blocking the EMT and Wnt/β-catenin pathway. Therefore, we surmise that miR-212 expression levels may be a useful indicator of NPC.
Materials and Methods
Clinical tissues
All experimental tissues were sampled from 33 patients with NPC at Heze Municipal Hospital and Linyi People’s Hospital. NPC patients who provided consent forms did not receive other treatments except for surgery. The study protocols and consent forms were approved by The Ethics Review Board of Anqiu People's Hospital (approval number 0015009). The surgically removed NPC or normal tissue was collected from the operation rooms, cut into small pieces (2×2×2 mm) and stored in -80° C before testing. Tissue sample was homogenized in Bullet Blender tissue homogenizer before RNA and protein isolation. For survival assay, we divided the 33 patients into two groups: low miR-212 expression group (n=15, >medium value) and high miR-212 expression group (n=18,
Cell lines
Normal nasopharyngeal epithelial cells NP69 (BNCC339267), NPC cell lines 6-10B (BNCC342441) and C666-1 (BNCC337872) were obtained from BeNa Culture Collection (BNCC, Beijing, China). The cells were maintained at 37° C in a humidified incubator with 5% CO2. RPMI-1640 medium containing 10% fetal bovine serum (FBS) was used for cell culture.
Transfection
miR-212 inhibitors, miR-212 mimics and the negative control miR sequence (NC), and TPD52 overexpression plasmid were purchased from ACON (Hangzhou, China). The miRNA negative control RNA sequence was based on common miRNA structure for use as negative experimental control in human and had been analyzed by BLAST against all human sequences and miRNA sequences in the current miRBase Database. siRNA against TPD52 was purchased from Santa Cruz (sc-45341). Transfections were performed in C666-1 cells using Lipofectamine 2000 (ThermoFisher).
Animal study
In vivo tumorigenesis assay was approved by the Animal Protection and Ethics Committee of the Anqiu People's Hospital. BALB/c nude mice (8-10 weeks) were from Beijing Weitonglihua company (Beijing, China). For Xenograft, control C666-1 cells and cells with miR-212 overexpression or inhibition (5 × 106) were suspended in normal saline (200 μl) and administrated subcutaneously. Tumor volumes were measured by using the formula: Volume (mm3) = (L x W x H) / 2, where L represents tumor length, W as tumor width, and H as tumor height. All mice in this study were euthanized by cervical traction 20 days following tumor cell inoculation.
qRT-PCR
RNA sample was extracted from C666-1 cells using Trizol reagent (Takara Bio, Dalian, China). cDNA was made via reverse transcription using the miScriptII RT kit (Qiagen, Shanghai, China). The cDNA solution, SYBR Green PCR Master Mix, and primers (Beijing Unique Biotechnology, Beijing, China) were mixed prior to pipetting into a 96-well plate. qRT-PCR was conducted using a 7300 System (Applied Biosystems, USA) and the relative expression of miR-212 and TPD52 were assessed using the 2-ΔΔCt method with U6 and GAPDH as controls, respectively. The primers were as follows: miR-212-F, CGCTAACAGTCTCCAGTC; miR-212-R, GTGCAGGGTCCGAGG T. U6-F, CGCAAGGATGACACGCAAATTC; U6-R, AACGCTTCACGAATTTGCGT; TPD52-F, AACAGAACATTGCCAAAGGGT G; TPD52-R, TGACTGAGCCAACAGACGAAA; GAPDH-F, AGCCACATCGCTCAGACAC; GAPDH-R, GCCCAATACGACCAAATCC.
Western blot
Cells were lysed in RIPA buffer (Beyotime, China) to isolate total proteins. The NE-PER nuclear extraction kit was used for nuclear proteins extraction according to the kit manual (ThermoFisher, 78833). Protein concentration was measured by BCA method and 15 μg was loaded and separated in a 10% SDS-PAGE. After electrophoresis, protein bands were transferred to PVDF membranes (Millipore, USA) and blocked with 5% non-fat milk in TBST. The membrane was probed with primary antibodies (4° C overnight) and secondary antibodies (RT 1 hour). Finally, the target proteins were visualized using the enhanced chemiluminescence (ECL) protein detection kit (Millipore, USA) and analyzed by Image-Pro Plus software version 6.0 (Media Cybernetics, USA). Primary antibodies of E-cadherin (1:1000, ab1416), TPD52 (1:1000, ab244523), N-cadherin (1:500, ab76011), Wnt1 (1:1000, ab15251), Vimentin (1:1000, ab92547), β-catenin (1:1000, ab32572), Cyclin D1 (1:500, ab16663), β-actin (1:3000, ab8226) and Lamin-B1 (1:1000, ab16048) were from abcam.
Immunohistochemistry
Paraffin sections of mice tissues were dewaxed in xylene and a series of descending ethanol solutions. 0.5% Triton X-100 was used for penetration and 50% goat serum was used for blocking. Sections were then incubated with Ki-67 or CD31 primary antibody overnight and developed with the DAB kit (Vector Laboratories, USA). Hematoxylin was used for counterstain and samples were dehydrated and mounted. The images were analyzed by Image-Pro Plus.
Methyl thiazolyl tetrazolium (MTT) assay
MTT cell proliferation and cytotoxicity assay kit (Solelybio #C0009, China) was used to compare the cell growth rate strictly according to the kit instruction. The absorbance at 490 nm was measured using a microplate reader (Molecular Devices).
Transwell
For invasion assay, the above chamber surface of the Transwell (8-μm pore size membranes) was pre-coated with Matrigel (BD Biosciences, USA). C666-1 cells (1×104 cells) were seeded in the above chamber, and the bottom one was filled with medium containing 20% FBS. After a 24-h incubation, the invaded cells were stained with 0.1% crystal violet and enumerated under a light microscope at 400× magnification. The Matrigel was not used in the migration assay, and other steps were the same as the invasion assay.
Wound healing
Cells were plated in 6-well plates until they formed a confluent monolayer. Cells were then scratched using a 100 μL pipette tip. The scratch wounds were captured using microscopy immediately after scratching and 24-48 h later.
Dual luciferase assay
The wildtype 3’-UTR of TPD52 containing the predicted miR-212 binding site (WT-TPD52) was mutated to disrupt the binding site through Q5 site-directed mutagenesis kit (NEB). The generated mutant 3’-UTR (Mut-TPD52) or WT-TPD52 was inserted into the pmirGLO dual-luciferase vector (Qcbio Science and Technologies Co., Ltd, Shanghai, China). MiR-212 mimics and the above vector were co-transfected into C666-1 cells. Dual-Glo Luciferase Assay System (Qcbio S & T, Shanghai, China) was used to measure luciferase activity 48 hours after transfection.
Statistical analysis
SPSS 13.0 (SPSS Inc., USA) and Graphpad Prism 6 (GraphPad, USA) were used for data analysis. Chi-squared Test, one-way analysis of variance followed by TUKEY post-hoc test were performed for multiple groups comparison. Student's t-test was performed between two groups. Survival curves were compared by Kaplan-Meier analysis with log-rank test. p<0.05 was considered as statistical significance.
Author Contributions
Bing Wang and Zheng Gong designed the experiments; Lili Sun performed the research; Juan Zheng analyzed the data; Jun Li and Xiao-Hui Gao wrote the manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This project was supported by our hospital.
References
- 1. Razak AR, Siu LL, Liu FF, Ito E, O’Sullivan B, Chan K. Nasopharyngeal carcinoma: the next challenges. Eur J Cancer. 2010; 46:1967–78. https://doi.org/10.1016/j.ejca.2010.04.004 [PubMed]
- 2. Du C, Ying H, Zhang Y, Huang Y, Zhai R, Hu C. Treatment for retropharyngeal metastatic undifferentiated squamous cell carcinoma from an unknown primary site: results of a prospective study with irradiation to nasopharyngeal mucosa plus bilateral neck. Oncotarget. 2017; 8:42372–81. https://doi.org/10.18632/oncotarget.16344 [PubMed]
- 3. Xu T, Ou X, Shen C, Hu C. Cetuximab in combination with chemoradiotherapy in the treatment of recurrent and/or metastatic nasopharyngeal carcinoma. Anticancer Drugs. 2016; 27:66–70. https://doi.org/10.1097/CAD.0000000000000294 [PubMed]
- 4. Nakahara S, Hanamoto A, Seo Y, Miyaguchi S, Yamamoto Y, Tomiyama Y, Yoshii T, Takenaka Y, Yoshioka Y, Isohashi F, Ogawa K, Inohara H. Chemoradiotherapy with weekly low-dose docetaxel and cisplatin concurrent with radiation for patients with locally advanced nasopharyngeal carcinoma, followed by adjuvant chemotherapy for selected patients. Jpn J Clin Oncol. 2016; 46:903–10. https://doi.org/10.1093/jjco/hyw100 [PubMed]
- 5. Shukla GC, Singh J, Barik S. MicroRNAs: Processing, Maturation, Target Recognition and Regulatory Functions. Mol Cell Pharmacol. 2011; 3:83–92. [PubMed]
- 6. Meng X, Wu J, Pan C, Wang H, Ying X, Zhou Y, Yu H, Zuo Y, Pan Z, Liu RY, Huang W. Genetic and epigenetic down-regulation of microRNA-212 promotes colorectal tumor metastasis via dysregulation of MnSOD. Gastroenterology. 2013; 145:426–36.e1. https://doi.org/10.1053/j.gastro.2013.04.004 [PubMed]
- 7. Xu L, Wang F, Xu XF, Mo WH, Xia YJ, Wan R, Wang XP, Guo CY. Down-regulation of miR-212 expression by DNA hypermethylation in human gastric cancer cells. Med Oncol. 2011 (Suppl 1); 28:S189–96. https://doi.org/10.1007/s12032-010-9691-0 [PubMed]
- 8. Incoronato M, Urso L, Portela A, Laukkanen MO, Soini Y, Quintavalle C, Keller S, Esteller M, Condorelli G. Epigenetic regulation of miR-212 expression in lung cancer. PLoS One. 2011; 6:e27722. https://doi.org/10.1371/journal.pone.0027722 [PubMed]
- 9. Liu J, Chen B, Yue B, Yang J. MicroRNA-212 suppresses the proliferation and migration of osteosarcoma cells by targeting forkhead box protein A1. Exp Ther Med. 2016; 12:4135–41. https://doi.org/10.3892/etm.2016.3880 [PubMed]
- 10. Li D, Li Z, Xiong J, Gong B, Zhang G, Cao C, Jie Z, Liu Y, Cao Y, Yan Y, Xiong H, Qiu L, Yang M, et al. MicroRNA-212 functions as an epigenetic-silenced tumor suppressor involving in tumor metastasis and invasion of gastric cancer through down-regulating PXN expression. Am J Cancer Res. 2015; 5:2980–97. [PubMed]
- 11. Wei LQ, Liang HT, Qin DC, Jin HF, Zhao Y, She MC. MiR-212 exerts suppressive effect on SKOV3 ovarian cancer cells through targeting HBEGF. Tumour Biol. 2014; 35:12427–34. https://doi.org/10.1007/s13277-014-2560-2 [PubMed]
- 12. Ma C, Nong K, Wu B, Dong B, Bai Y, Zhu H, Wang W, Huang X, Yuan Z, Ai K. miR-212 promotes pancreatic cancer cell growth and invasion by targeting the hedgehog signaling pathway receptor patched-1. J Exp Clin Cancer Res. 2014; 33:54. https://doi.org/10.1186/1756-9966-33-54 [PubMed]
- 13. Shi B, Wang Y, Yin F. MALAT1/miR-124/Capn4 axis regulates proliferation, invasion and EMT in nasopharyngeal carcinoma cells. Cancer Biol Ther. 2017; 18:792–800. https://doi.org/10.1080/15384047.2017.1373214 [PubMed]
- 14. Mrozik KM, Blaschuk OW, Cheong CM, Zannettino AC, Vandyke K. N-cadherin in cancer metastasis, its emerging role in haematological malignancies and potential as a therapeutic target in cancer. BMC Cancer. 2018; 18:939. https://doi.org/10.1186/s12885-018-4845-0 [PubMed]
- 15. Satelli A, Li S. Vimentin in cancer and its potential as a molecular target for cancer therapy. Cell Mol Life Sci. 2011; 68:3033–46. https://doi.org/10.1007/s00018-011-0735-1 [PubMed]
- 16. Lin YC, You L, Xu Z, He B, Mikami I, Thung E, Chou J, Kuchenbecker K, Kim J, Raz D, Yang CT, Chen JK, Jablons DM. Wnt signaling activation and WIF-1 silencing in nasopharyngeal cancer cell lines. Biochem Biophys Res Commun. 2006; 341:635–40. https://doi.org/10.1016/j.bbrc.2005.12.220 [PubMed]
- 17. Fu W, Tao T, Qi M, Wang L, Hu J, Li X, Xing N, Du R, Han B. MicroRNA-132/212 Upregulation Inhibits TGF-β-Mediated Epithelial-Mesenchymal Transition of Prostate Cancer Cells by Targeting SOX4. Prostate. 2016; 76:1560–70. https://doi.org/10.1002/pros.23241 [PubMed]
- 18. Jia P, Wei G, Zhou C, Gao Q, Wu Y, Sun X, Li X. Upregulation of MiR-212 Inhibits Migration and Tumorigenicity and Inactivates Wnt/β-Catenin Signaling in Human Hepatocellular Carcinoma. Technol Cancer Res Treat. 2018; 17:1533034618765221. https://doi.org/10.1177/1533034618765221 [PubMed]
- 19. Wang R, Xu J, Saramäki O, Visakorpi T, Sutherland WM, Zhou J, Sen B, Lim SD, Mabjeesh N, Amin M, Dong JT, Petros JA, Nelson PS, et al. PrLZ, a novel prostate-specific and androgen-responsive gene of the TPD52 family, amplified in chromosome 8q21.1 and overexpressed in human prostate cancer. Cancer Res. 2004; 64:1589–94. https://doi.org/10.1158/0008-5472.can-03-3331 [PubMed]
- 20. Tennstedt P, Bölch C, Strobel G, Minner S, Burkhardt L, Grob T, Masser S, Sauter G, Schlomm T, Simon R. Patterns of TPD52 overexpression in multiple human solid tumor types analyzed by quantitative PCR. Int J Oncol. 2014; 44:609–15. https://doi.org/10.3892/ijo.2013.2200 [PubMed]
- 21. Byrne JA, Maleki S, Hardy JR, Gloss BS, Murali R, Scurry JP, Fanayan S, Emmanuel C, Hacker NF, Sutherland RL, Defazio A, O’Brien PM. MAL2 and tumor protein D52 (TPD52) are frequently overexpressed in ovarian carcinoma, but differentially associated with histological subtype and patient outcome. BMC Cancer. 2010; 10:497. https://doi.org/10.1186/1471-2407-10-497 [PubMed]
- 22. Byrne JA, Balleine RL, Schoenberg Fejzo M, Mercieca J, Chiew YE, Livnat Y, St Heaps L, Peters GB, Byth K, Karlan BY, Slamon DJ, Harnett P, Defazio A. Tumor protein D52 (TPD52) is overexpressed and a gene amplification target in ovarian cancer. Int J Cancer. 2005; 117:1049–54. https://doi.org/10.1002/ijc.21250 [PubMed]
- 23. Kumamoto T, Seki N, Mataki H, Mizuno K, Kamikawaji K, Samukawa T, Koshizuka K, Goto Y, Inoue H. Regulation of TPD52 by antitumor microRNA-218 suppresses cancer cell migration and invasion in lung squamous cell carcinoma. Int J Oncol. 2016; 49:1870–80. https://doi.org/10.3892/ijo.2016.3690 [PubMed]
- 24. Zhang HQ, Sun Y, Li JQ, Huang LM, Tan SS, Yang FY, Li H. The expression of microRNA-324-3p as a tumor suppressor in nasopharyngeal carcinoma and its clinical significance. Onco Targets Ther. 2017; 10:4935–43. https://doi.org/10.2147/OTT.S144223 [PubMed]
- 25. Jiping Z, Ming F, Lixiang W, Xiuming L, Yuqun S, Han Y, Zhifang L, Yundong S, Shili L, Chunyan C, Jihui J. MicroRNA-212 inhibits proliferation of gastric cancer by directly repressing retinoblastoma binding protein 2. J Cell Biochem. 2013; 114:2666–72. https://doi.org/10.1002/jcb.24613 [PubMed]
- 26. Gu C, Wang Z, Jin Z, Li G, Kou Y, Jia Z, Yang J, Tian F. MicroRNA-212 inhibits the proliferation, migration and invasion of renal cell carcinoma by targeting X-linked inhibitor of apoptosis protein (XIAP). Oncotarget. 2017; 8:92119–33. https://doi.org/10.18632/oncotarget.20786 [PubMed]
- 27. Lv ZD, Yang DX, Liu XP, Jin LY, Wang XG, Yang ZC, Liu D, Zhao JJ, Kong B, Li FN, Wang HB. MiR-212-5p Suppresses the Epithelial-Mesenchymal Transition in Triple-Negative Breast Cancer by Targeting Prrx2. Cell Physiol Biochem. 2017; 44:1785–95. https://doi.org/10.1159/000485785 [PubMed]
- 28. Luo XJ, Tang DG, Gao TL, Zhang YL, Wang M, Quan ZX, Chen J. MicroRNA-212 inhibits osteosarcoma cells proliferation and invasion by down-regulation of Sox4. Cell Physiol Biochem. 2014; 34:2180–8. https://doi.org/10.1159/000369661 [PubMed]
- 29. Liu H, Li C, Shen C, Yin F, Wang K, Liu Y, Zheng B, Zhang W, Hou X, Chen X, Wu J, Wang X, Zhong C, et al. MiR-212-3p inhibits glioblastoma cell proliferation by targeting SGK3. J Neurooncol. 2015; 122:431–9. https://doi.org/10.1007/s11060-015-1736-y [PubMed]
- 30. Zhang Y, Li Y, Wang J, Lei P. Long non-coding RNA ferritin heavy polypeptide 1 pseudogene 3 controls glioma cell proliferation and apoptosis via regulation of the microRNA-224-5p/tumor protein D52 axis. Mol Med Rep. 2018; 18:4239–46. https://doi.org/10.3892/mmr.2018.9491 [PubMed]
- 31. Chen H, Xu H, Meng YG, Zhang Y, Chen JY, Wei XN. miR-139-5p regulates proliferation, apoptosis, and cell cycle of uterine leiomyoma cells by targeting TPD52. Onco Targets Ther. 2016; 9:6151–60. https://doi.org/10.2147/OTT.S108890 [PubMed]
- 32. Zhang Z, Wang J, Gao R, Yang X, Zhang Y, Li J, Zhang J, Zhao X, Xi C, Lu X. Downregulation of MicroRNA-449 Promotes Migration and Invasion of Breast Cancer Cells by Targeting Tumor Protein D52 (TPD52). Oncol Res. 2017; 25:753–61. https://doi.org/10.3727/096504016X14772342320617 [PubMed]
- 33. Li G, Yao L, Zhang J, Li X, Dang S, Zeng K, Zhou Y, Gao F. Tumor-suppressive microRNA-34a inhibits breast cancer cell migration and invasion via targeting oncogenic TPD52. Tumour Biol. 2016; 37:7481–91. https://doi.org/10.1007/s13277-015-4623-4 [PubMed]
- 34. Zhao X, Chu J. MicroRNA-379 suppresses cell proliferation, migration and invasion in nasopharyngeal carcinoma by targeting tumor protein D52. Exp Ther Med. 2018; 16:1232–40. https://doi.org/10.3892/etm.2018.6302 [PubMed]
- 35. Yin W, Shi L, Mao Y. MicroRNA-449b-5p suppresses cell proliferation, migration and invasion by targeting TPD52 in nasopharyngeal carcinoma. J Biochem. 2019; 166:433–40. https://doi.org/10.1093/jb/mvz057 [PubMed]
- 36. Ji Y, Wang M, Li X, Cui F. The Long Noncoding RNA NEAT1 Targets miR-34a-5p and Drives Nasopharyngeal Carcinoma Progression via Wnt/β-Catenin Signaling. Yonsei Med J. 2019; 60:336–45. https://doi.org/10.3349/ymj.2019.60.4.336 [PubMed]
- 37. Teufel A, Marquardt JU, Galle PR. Next generation sequencing of HCC from European and Asian HCC cohorts. Back to p53 and Wnt/β-catenin. J Hepatol. 2013; 58:622–4. https://doi.org/10.1016/j.jhep.2012.10.006 [PubMed]