



Introduction
Aging is universally characterized by a progressive loss of physiological integrity, leading to organ failure [1, 2]. This deterioration is the principal risk factor for numerous chronic diseases, such as type 2 diabetes, cancer, cardiovascular disorders, and neurodegenerative diseases [3]. Therefore, interventions to prevent or attenuate age-related degeneration are an unmet need. To date, some drugs that can affect lifespan and healthspan have been found, such as rapamycin and metformin, which increase lifespan in multiple species [4, 5]. Both drugs have been proven to be relatively safe over decades of clinical use; however, side effects are sometimes observed. Therefore, the discovery of new anti-aging drugs is urgent. Endogenous metabolites that can regulate lifespan have considerable advantages because they have limited toxicity and side effects. Recently, several metabolites have been identified to modulate aging. For example, one study found that dietary supplementation with the nematode eicosapentaenoyl ethanolamide resulted in a significant reduction in lifespan and stress tolerance [6]. Another study reported that oxaloacetate supplementation increased lifespan in a DAF-16-dependent manner in C. elegans [7]. Additionally, a previous study also reported that dietary supplementation with the metabolite α-ketoglutarate lengthened lifespan by inhibiting ATP synthase and TOR signaling [8].
Intermediates of pyrimidine metabolism are essential biomolecules, as they participate in diverse cellular functions, such as the synthesis of DNA, RNA, lipids, and carbohydrates [9]. Previous studies have reported that pyrimidine metabolism disruption causes some diseases, such as Alzheimer's disease, immunodeficiency, and growth retardation [10, 11], and the downregulation of pyrimidine metabolism was linked to aging in mice [12]. In our previous study, we reported that the levels of pyrimidine intermediates were significantly decreased in aged worms, suggesting that pyrimidine metabolism is related to the aging of C. elegans [13]. However, the mechanism by which the pyrimidine metabolism pathway regulates longevity remains unknown.
In this study, we used C. elegans as a model organism to explore the mechanism by which pyrimidine metabolism regulates aging. C. elegans is an excellent model for studying the molecular mechanism of longevity modulation. Genetic studies in C. elegans led to the identification of numerous molecular pathways that regulate longevity, including the insulin/insulin-like growth factor (IGF), target of rapamycin (TOR), germline signaling pathways, proteostasis, and mitochondrial function pathways [14–17]. These genetic pathways identified to regulate longevity are evolutionarily conserved, from yeast to human. Here, we characterized the biological effects of intermediates of the pyrimidine pathway, including thymine, orotate, β-aminoisobutyrate, uridine and cytidine. We found that pyrimidine intermediates could significantly prolong the lifespan of C. elegans through the inhibition of reproductive signaling, the activation of the nuclear receptors NHR-49 and DAF-12, and the upregulation of the transcription factor DAF-16.
Results
Intermediate metabolites in the pyrimidine metabolism pathway increase the lifespan of C. elegans
To dissect the mechanism by which the pyrimidine metabolism pathway (Figure 1A)regulates lifespan, we detected the effect of intermediate metabolites in this metabolic pathway on the lifespan of C. elegans by feeding worms these endogenous small molecules. We found that feeding animalsthymine, β-aminoisobutyrate, orotate, uridine and cytidine at a concentration of 0.5 mM on Escherichia. coli OP50, which were heat-inactivated to avoid the catalysis of compounds by bacteria, could significantly extend lifespan by 15.2, 9.69, 14.8, 9.83, 7.87%, respectively, compared with the nontreated control (Figure 1B–1F). By contrast, treatment with uracil and thymidine at the same concentration had no effect on lifespan (Figure 1G–1H). These data suggest that the pyrimidine metabolism pathway plays an important role in regulating the lifespan of C. elegans. We also found that worms treated with 0.5 mM uridine and cytidine exhibited 5-fluro-2′-deoxyuridine (FUdR) resistance, as 10 μM FUdR could not prevent self-fertilization. Therefore, in all subsequent lifespan analyses using uridine and cytidine, animals were maintained on the plates without FUdR.
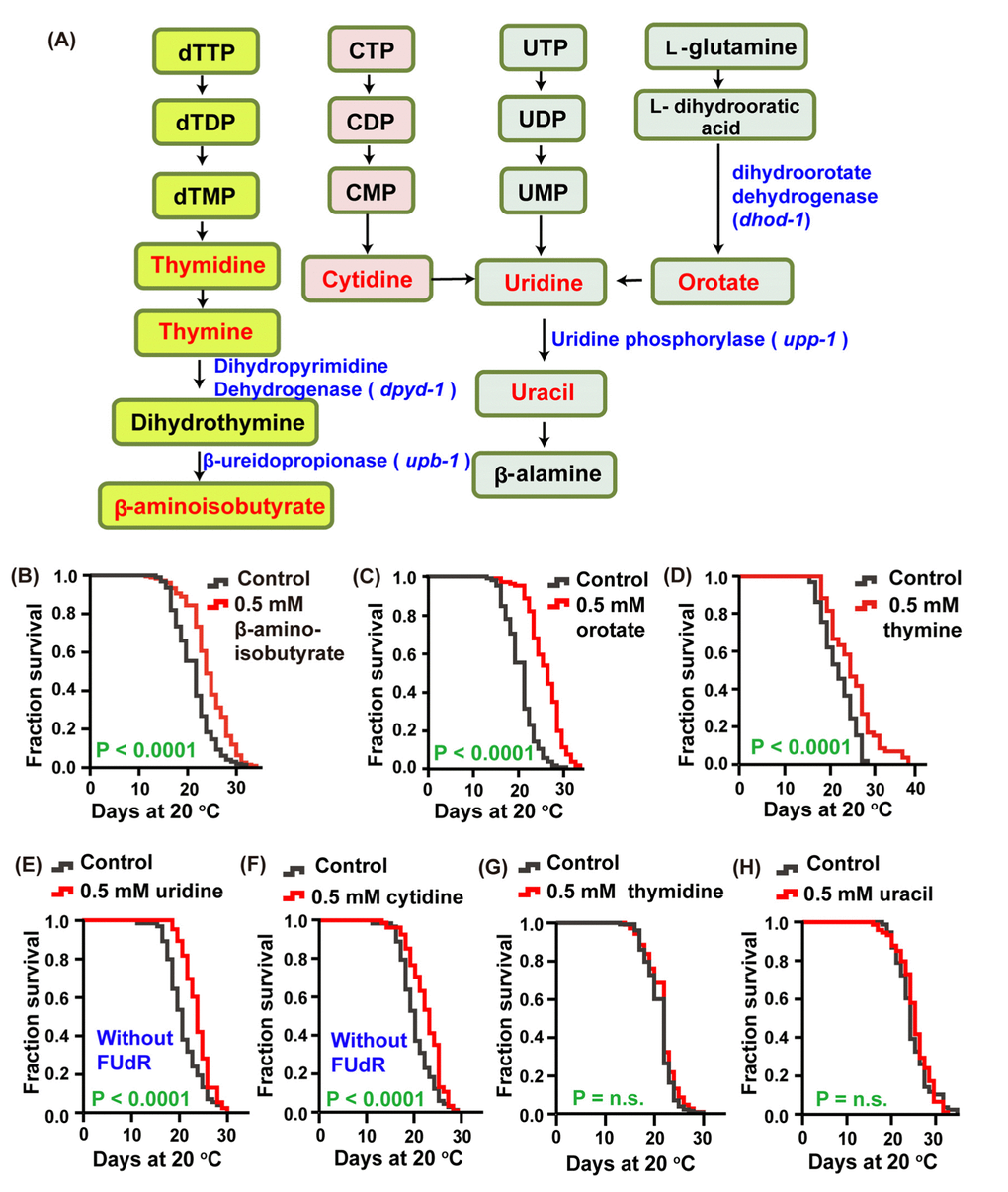
Figure 1. The intermediate metabolites of the pyrimidine metabolism pathway extended the lifespan of C. elegans. (A) Schematic representation of the pyrimidine metabolism pathway. (B–H) Lifespan of worms treated with 0.5 mM (B) β-aminoisobutyrate (red), (C) orotate (red), (D) thymine (red), (E) uridine (red), (F) cytidine (red), (G) thymidine (red), (H) uracil (red), and vehicle (gray) at 20 °C. The treatments were administered beginning on day 1 of adulthood. Lifespan was analyzed using the Kaplan-Meier test, and P values were calculated using the log-rank test; no significant difference was abbreviated as n.s. Data are representative of at least two independent experiments, and lifespan values are listed in Supplementary Table 1.
Essential genes in the pyrimidine metabolic pathway regulate the aging of C. elegans
In C. elegans, dpyd-1 encodes the C. elegans ortholog of human dihydropyrimidine dehydrogenase (DPYD), which is the first and rate-limiting enzyme for the metabolism of thymine to 5,6-dihydrothymine [18]. We detected that knockdown of the dpyd-1 gene by RNAi, which dramatically decreased the mRNA level, extended the lifespan of worms (Figure 2A and 2D).
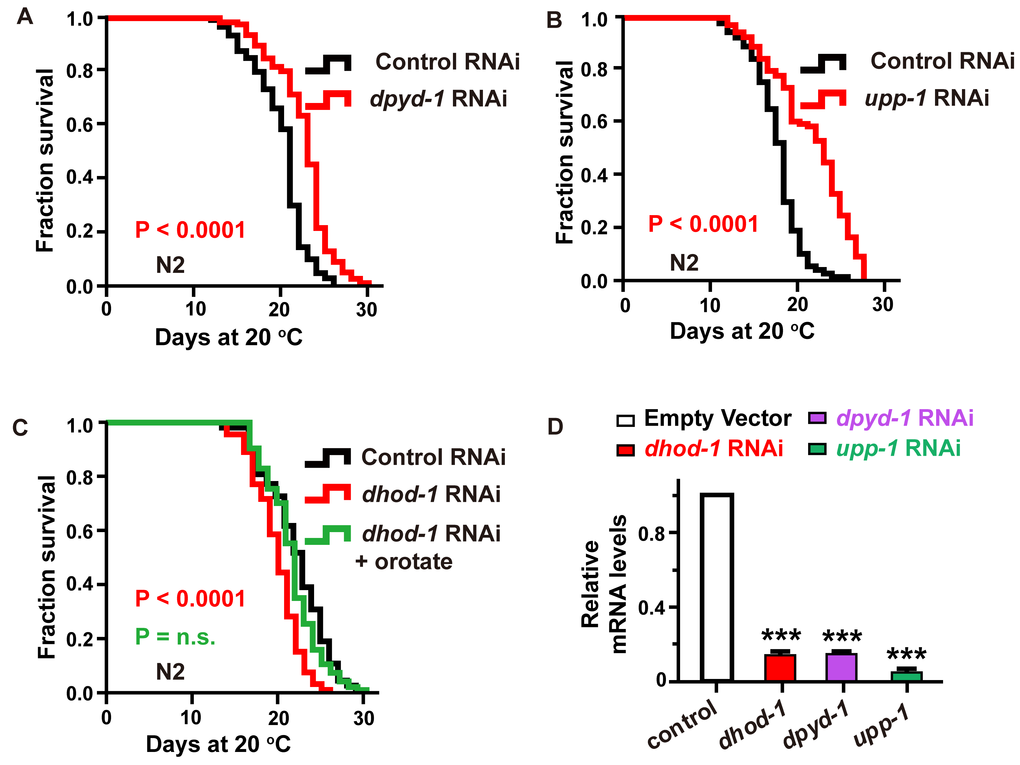
Figure 2. Characterization of the genes involved in the pyrimidine metabolism pathway as aging-related genes. (A–C) The effect of (A) dpyd-1 RNAi (red) and (B) upp-1 RNAi (red) on lifespan. (C) Lifespan of wild-type N2 exposed to control, dhod-1 RNAi, and dhod-1 RNAi with orotate supplementation. Lifespan values of repetitions are listed in Supplementary Table 1. (D) dhod-1, dpyd-1, and upp-1 RNA levels in whole worms after treatment of C. elegans with RNAi against dhod-1, dpyd-1 and upp-1, respectively, versus control RNAi. (mean ± SD, n=3, *** P<0.001, Student’s t test).
Similarly, the C. elegans UP homologous protein (UPP-1) exhibits both uridine and thymidine phosphorylase activity [19]. We found that lifespan was significantly extended (Figure 2B and 2D) following the exposure of C. elegans to upp-1 RNAi bacteria.
Furthermore, the C. elegans DHODH (dihydroorotate dehydrogenase (quinone)) homologous protein DHOD-1 catalyzed the transition of dihydroorotate to orotate [20]. We observed that the lifespan of C. elegans was significantly decreased following exposure to dhod-1 RNAi (Figure 2C and 2D). Additionally, we found that the decreased lifespan caused by dhod-1 impairment could be rescued by supplementation with 2 mM orotate (Figure 2C). In previous experiments, metabolites were applied at a concentration of 0.5 mM when feeding heat-inactivated OP50. However, for RNAi experiments, in which bacteria cannot be inactivated, 2 mM was used to compensate for the loss caused by the portion of treatments metabolized by live HT115 bacteria.
However, the RNAi knockdown of upb-1, a gene that catalyzes the transition of dihydrothymine to β-aminoisobutyrate [21], had no effect on the lifespan of treated worms (Supplementary Figure 1A and 1B, Supporting information). One possibility is that upb-1 is involved in several metabolic pathways, and its loss of function may cause some metabolite variations instead of exclusive changes in the levels of β-aminoisobutyrate. Accordingly, these results were consistent with a direct effect of metabolites, including thymine, uridine, cytidine, and orotate, on longevity.
Thymine regulates aging via a mechanism dependent on the transcription factor FOXO/DAF-16 and the nuclear receptors DAF-12 and NHR-49
Thymine is a representative intermediate metabolite in the pyrimidine metabolism pathway, and it is a stable molecule that had the best effect on lifespan extension among all the metabolites that benefited the longevity of C. elegans. Therefore, we chose thymine to systematically explore the molecular mechanism of the pyrimidine pathway. We exposed wild-type N2 worms to different concentrations of thymine and found that animals raised on plates containing 2 mM of thymine showed the largest lifespan extension of 18.3% (Figure 3A). Worms exposed to 10 mM and 0.5 mM also had moderately extended the lifespan (Figure 3A). At the lower concentrations of 0.1 and 0.05 mM, no lifespan extension was observed (Figure 3A). Therefore, thymine was applied at a concentration of 2 mM in all subsequent experiments.
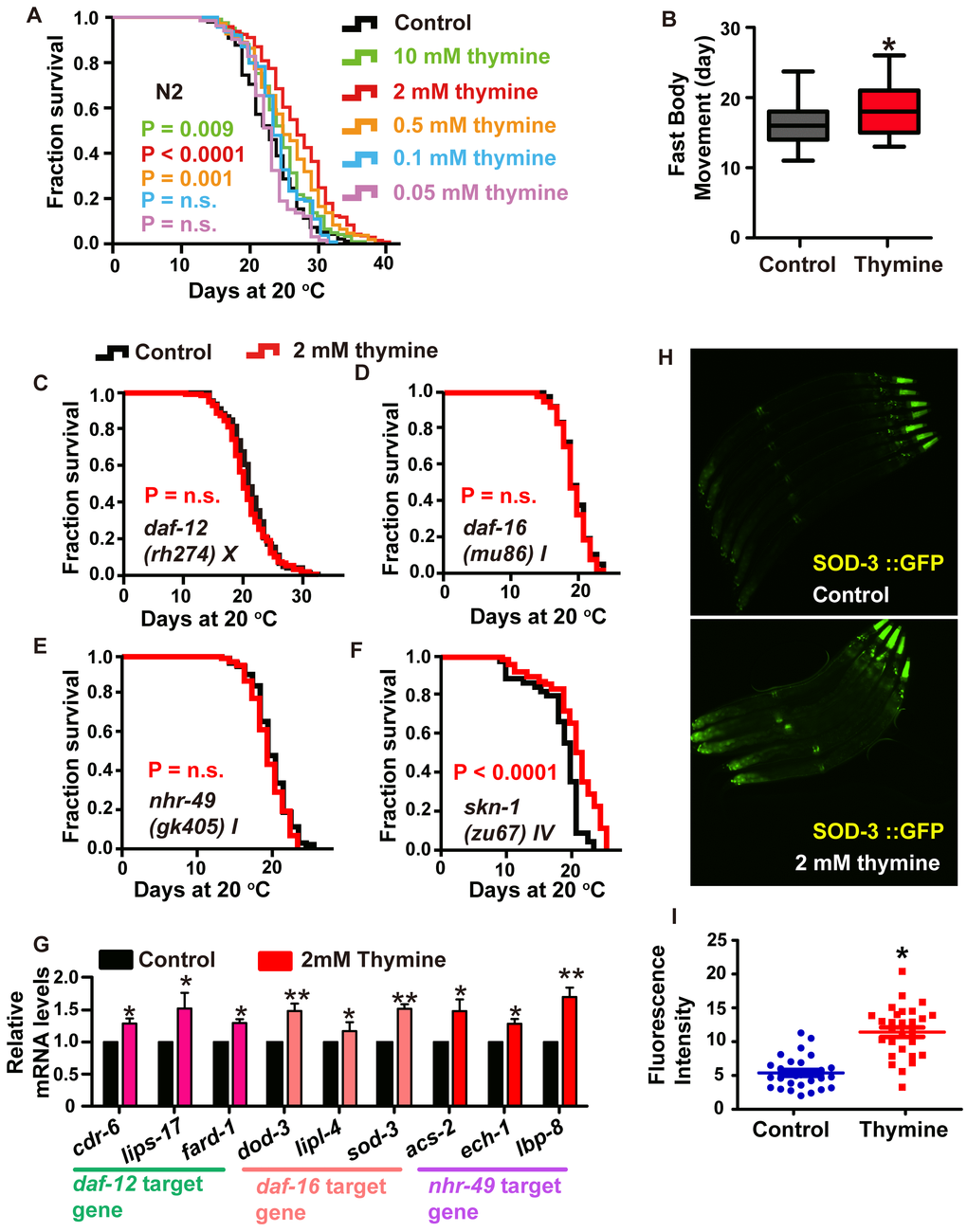
Figure 3. Lifespan extension induced by thymine treatment is mediated by the nuclear receptors DAF-12 and NHR-49, and the transcription factor DAF-16/FOXO. (A) Effects of thymine supplementation at increasing concentrations (0.05-10 mM) (P value determined by log-rank tests). (B) Age-related movements of worms treated with 2 mM thymine and vehicles. Data are the mean ± SD; * P <0.05 (Student’s t test). (C–F) Lifespan analysis in (C) daf-12 (rh274), (D) daf-16 (mu86), (E) nhr-49 (gk405), and (F) skn-1 (zu67) worms treated with 2 mM thymine (red) and vehicle (black), on heat-inactivated E. coli OP50 (P value by log-rank test). Lifespan values of replicated experiments are summarized in Supplementary Table 1. (G) QPCR analysis of the mRNA level of target genes of daf-12, daf-16 and nhr-49 in C. elegans treated with 2 mM thymine versus control. Data are the mean ± SD, n=3, * p<0.05 (Student’s t test). (H–I) Images and quantification of GFP fluorescence. *p < 0.05; n=30 (Student’s t test).
To assess whether healthspan was affected, we measured an age-related phenotype, body movement, which is one of the most obvious behavioral abnormalities associated with nematode aging [22]. We analyzed the body movement of animals treated with or without 2 mM thymine. Our results showed that body movement progressively declined during aging, while the decrease in body movement with aging was delayed by thymine (Figure 3B).
To explore whether and to what extent known transcriptional modulators of lifespan might contribute to the longevity phenotype caused by thymine, we performed survival analyses of different mutants. SKN-1, a homolog of mammalian Nrf2 (nuclear factor-erythroid related factor 2) [23], appeared dispensable for thymine-mediated lifespan extension, as thymine could further increase the lifespan of the skn-1 mutant; moreover, the expression of a target gene of skn-1 (gst-4) and the Pgst-4::GFP reporter were not significantly different between animals treated with or without 2 mM thymine. (Figure 3F and Supplementary Figure 2). The C. elegans DAF-16 [24], nuclear receptor PPARα homolog NHR-49 [25] and steroid nuclear receptor DAF-12 [26] appeared to be involved since the impairment of nhr-49, daf-12 and daf-16 abolished the lifespan extension induced by thymine (Figure 3C–3E). In addition, several major target genes of daf-16 (sod-3, dod-3 and lipl-4) [23, 27], daf-12 (card-6, lips-17 and fard-1) [28] and nhr-49 (acs-2, ech-1 and lbp-8) [29] were significantly increased when worms were exposed to 2 mM thymine (Figure 3G). Additionally, the expression of the Psod-3::GFP reporter in transgenic worms that were exposed to 2 mM thymine was significantly higher than that in nonexposed worms (Figure 3H and 3I). Taken together, these results indicated that thymine-induced lifespan extension requires the function of DAF-12, NHR-49, and DAF-16.
The effect of thymine on lifespan extension may be conferred by a reduction in germline signaling
We next dissected which longevity pathway is required for the lifespan extension induced by thymine through testing its effects in corresponding mutants. The ability of the Notch signaling pathway to sense nucleotide abundance is reminiscent of glp-1 [30], which encodes a Notch family receptor and is essential for the mitotic proliferation of germline cells [31]. The glp-1 loss-of function (lf) mutants have obviously prolonged lifespan when maintained at the nonpermissive temperature due to failed germline proliferation, which is dependent on the transcriptional activities of DAF-16, DAF-12, and NHR-49 [16, 32, 33]. Therefore, we first tested whether the longevity benefit of thymine was mediated by the Notch signaling pathway. Treatment with 2 mM thymine failed to further increase the lifespan of glp-1 mutants (Figure 4A), suggesting that thymine might regulate the germline signal to extend lifespan. The downregulation of germline signaling extends lifespan, usually accompanied by a decrease in the production of progeny [34]. However, we did not find that thymine decreased the daily progeny production, total progeny and germ cells of each worm (Figure 4E–4H). One possible explanation is that thymine does inhibit reproductive signals, but this inhibition does not reach a significant extent to affect the amount of eggs laid. Reduced reproductive signals is usually accompanied by a change in lipid metabolism [35]. To determine whether thymine affects fat metabolism, we used Oil-Red-O (ORO) staining. We found that wild-type animals treated with 2 mM thymine had significantly increased ORO staining (Figure 4I).
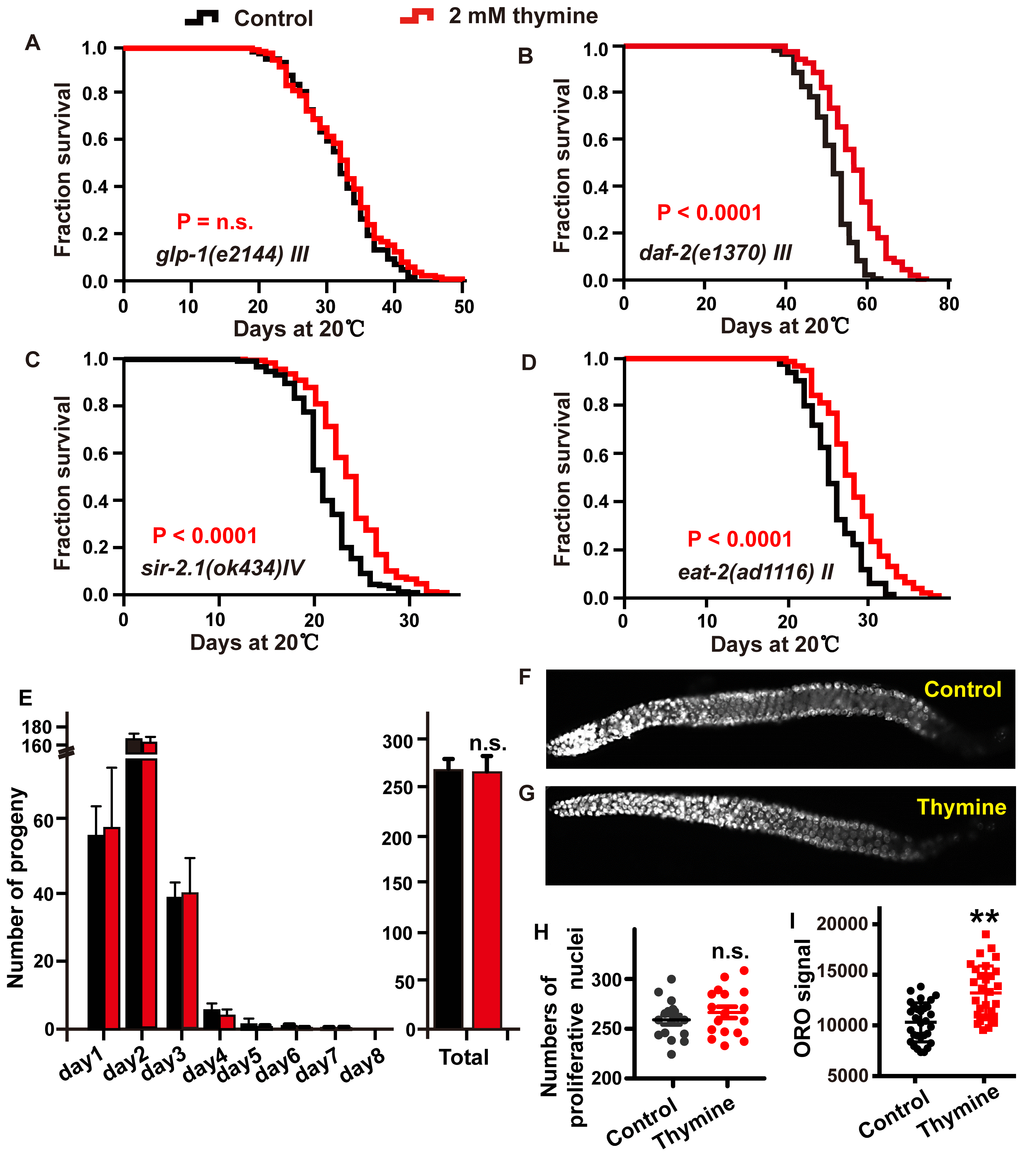
Figure 4. The lifespan extension induced by thymine depends on reproductive signals. (A–D) Lifespan analysis of (A) glp-1 (e2144), (B) daf-2 (e1370), (C) sir-2.1 (ok434), and (D) eat-2 (ad1116) animals treated with 2 mM thymine (red) and vehicle (black), on heat-inactivated E. coli OP50 (log-rank test). Lifespan values of replicated experiments are summarized in Supplementary Table 1. (E) The number of daily progeny and the total number of progeny of wild-type N2 worms treated with 2 mM thymine or vehicle. (F, G) DAPI-stained image of worm gonads. (H) Quantification of germline stem cells in wild-type N2 animals treated with 2 mM thymine or vehicle; n.s. not significant (Student’s t test). (I) Quantification of ORO staining. Mean ± SD.; n ≥ 30 per condition; P value was calculated using Student’s test; no significance is abbreviated as n.s. *p < 0.05, **p < 0.01, ***p < 0.001.
DAF-16 is downstream of several signaling pathways in addition to the reproduction signaling pathway, such as the IGF signaling pathway [24]. To explore whether thymine contributes to lifespan extension by influencing the IIS signaling pathway in a DAF-16-dependent manner, we detected the effect of thymine on long-lived mutants of the insulin-like receptor daf-2. We observed that the lifespan of daf-2 was significantly increased by thymine treatment, suggesting that the lifespan extension induced by thymine might not depend on the IIS pathway (Figure 4B).
The activation of DAF-16 by thymine was reminiscent of the silent information regulator 2 (SIR2), which encodes a nicotinamide adenine dinucleotide (NAD)-dependent deacetylase. SIR-2.1 could bind to DAF-16 and extend lifespan in a 14-3-3 protein-dependent manner [36]. We investigated whether SIR-2.1 mediated the lifespan extension caused by thymine using a null mutant sir-2.1 strain. Thymine extended the lifespan of the sir-2.1 mutant, indicating that SIR-2.1 was dispensable for thymine-mediated lifespan extension (Figure 4C).
Dietary restriction (DR) plays a significant role in the aging process in a wide range of species. The nuclear receptor NHR-49 and the transcription factor DAF-16 [37, 38], which function as mediators of the lifespan extension induced by thymine, were activated to respond to low energy and modulate lifespan under starvation conditions. Therefore, to determine whether the ability of thymine to extend lifespan was related to an effect similar to DR, we used a pharyngeal pumping defective eat-2 mutant, which was considered DR-constitutive due to its reduced food intake [39]. As shown in Figure 4D, thymine treatment significantly increased the mean lifespan of the eat-2 mutant, indicating that thymine acted through mechanisms different from the DR related mechanisms to extend the lifespan of C. elegans.
Other pyrimidine intermediates extend the lifespan of C. elegans in a similar mechanism mediated by thymine
In addition to thymine, other metabolites of the pyrimidine metabolism pathway, including β-aminoisobutyrate, uridine, cytidine and orotate could promote the lifespan of C. elegans. We tested whether the mechanism contributing to thymine-induced lifespan regulation was also involved in the lifespan extension induced by other pyrimidine intermediates. Our results revealed that β-aminoisobutyrate, uridine, cytidine, and orotate failed to further increase the lifespan of glp-1 (Figure 5A–5D) and daf-12 (Figure 6) mutants, which represent the reproduction signaling pathway. Similarly, the lipid content of wild-type worms was significantly increased when animals were exposed to 0.5 mM β-aminoisobutyrate, uridine, cytidine, and orotate (Figure 5E). Moreover, similar to the effect of thymine on reproduction, these metabolites could not affect germline stem cells of C. elegans (Figure 5F and 5G). Collectively, these results demonstrated that pyrimidine intermediates increased the lifespan of C. elegans by influencing the reproduction signaling pathway rather than by directly influencing fecundity.

Figure 5. Intermediate metabolites in pyrimidine metabolism influence the lifespan of C. elegans through regulating reproductive signals. (A–D) Lifespan analysis of glp-1 (e2144) animals treated with 0.5 mM (A) β-aminoisobutyrate (red), (B) cytidine (red), (C) orotate (red), (D) uridine (red), and water (black) on heat-inactivated E. coli OP50 (P value by log-rank test). The replicated data are summarized in Supplementary Table 1. (E) Quantification of ORO staining. Mean ± SD.; n ≥ 30 (Student’s t test). (F) DAPI-stained image of the gonads of N2 worms treated with 0.5 mM β-aminoisobutyrate, cytidine, orotate, and uridine and the untreated control. (G) Quantification of the germline stem cells in wild-type N2 worms treated with or without 0.5 mM of the metabolites. P values were calculated using Student’s t test.
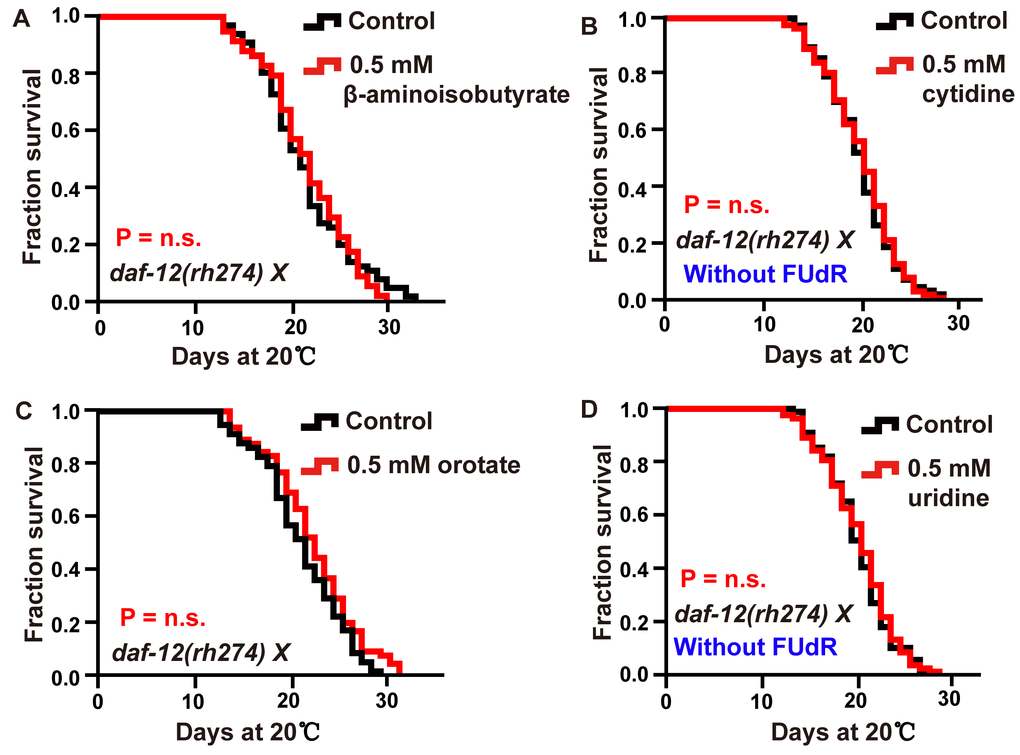
Figure 6. Intermediate metabolites in pyrimidine metabolism extended adult lifespan in a DAF-12-dependent manner. (A–D) Lifespan of daf-12 (rh274) animals treated with 0.5 mM (A) β-aminoisobutyrate, (B) cytidine (red), (C) orotate (red), (D) uridine (red), and vehicle (black). The P value was calculated by the log-rank test. Replicates of these experiments and statistical details are summarized in Supplementary Table 1.
Discussion
Pyrimidine metabolism has been broadly studied in many organisms. The importance of this metabolic pathway is due to the fact that they provide pyrimidine nucleosides, which are essential components of many biomolecules. Additionally, pyrimidine metabolism disorders can cause some diseases, such as orotic aciduria, which results from uridine monophosphate synthase (UMPS) deficiency [40]. Previously, a large number of studies focused on the relationship between pyrimidine and cancer. Pyrimidine derivatives have been used as anticancer agents, such as 5-fluorouracil (5-FU), which is a predrug molecule and is converted to an active drug via the pyrimidine biosynthesis pathway. Therefore, the function of this drug is dependent on the activities of the pyrimidine synthesis enzymes [41]. Moreover, our previous study suggested that pyrimidine metabolism is closely associated with aging of C. elegans [13]. In the present study, we screened some intermediates involved in pyrimidine metabolism and found that thymine, uridine, cytidine, orotate, and β-aminoisobutyrate could significantly increase the lifespan of C. elegans (Figure 1B–1F). In addition, we demonstrated an uncharacterized mechanism by which pyrimidine nucleotides could extend the lifespan of C. elegans by inhibiting the Notch-associated reproduction signaling pathway. These result is consistent with the analysis of concentration of pyrimidine intermediates between glp-1 mutants and wild-type in our previous study. In that study, we found that there are many difference in concentration of pyrimidine intermediates between glp-1 mutant and wild-type N2, such as β-aminoisobutyrate, uracil, UMP and CDP and so on [13]. However, due to the limitation of detection sensitivity, we did not detect changes in concentration of thymine, thymidine and uridine in previous study. Here, we further performed the analysis of levels of pyrimidine intermediates between glp-1 mutant and wild-type N2, and found that the level of thymine and thymidine significantly increased in glp-1 mutant compared with wild-type N2, those of uridine moderately increased but no significant difference (Supplementary Figure 4). In the in-depth pathway analysis, we revealed that these metabolites extend lifespan by downregulating reproductive signals and then evoking the nuclear receptors DAF-12 and NHR-49, and the transcription factor DAF-16.
Nucleotide homeostasis is tightly associated with human health. In humans, abnormally elevated concentrations of pyrimidine intermediate metabolites can lead to a high incidence of many diseases. For example, a high nucleotide supply could lead to the uncontrolled proliferation of cancer cells [42]. Increased uracil levels will result in urea cycle disorders [43]. The depletion of 5-phosphoribosyl-1-pyrophosphate (PRPP) causes orotate accumulation, which can result in secondary orotic aciduria [44]. However, pyrimidine nucleotide starvation has also been linked to some diseases, such as developmental delay, seizures, alopecia and recurrent infections [45]. In our study, we found that the elevated concentrations of pyrimidine metabolism intermediates induced by exogenous supplementation could significantly increase lifespan. The mechanism underlying this complicated regulatory feedback loop in worms that modulates internal nucleotide homeostasis remains to be explained. Based on a previous study [30], worms sense nucleotide levels through the Notch signaling pathway. When the level of nucleotides was low, animals shut down germline proliferation to protect both the mother and the progeny from irreversible and deleterious effects resulting from the exhaustion of the limited nucleotide pool. When the nucleic acid level was slightly increased but the toxic levels were not reached, animals showed a slight inhibition of reproductive signals and achieved lifespan extension. Certainly, the abnormal elevation of nucleotides could lead to the uncontrolled proliferation of cells, as previously mentioned [42]. These results indicated that as a nucleic acid receptor, the Notch signaling pathway could be downregulated under low nucleotide conditions, while under high nucleotide conditions, a negative feedback loop is activated to downregulate the Notch signaling pathway, ultimately maintaining nucleic acid homeostasis to maintain a normal physiological state (Supplemenary Figure 5).
UP is a member of the pyrimidine biosynthesis family of enzymes and is a key regulator of uridine homeostasis. The C. elegans UP homologs protein (UPP-1) exhibited both uridine and thymidine phosphorylase activity in vitro. UP activity is typically upregulated in various tumor tissues [46]. A previous study found that the lifespan of upp-1 mutant worms was reduced by approximately 30% compared with that of wild-type worms [47]. However, in contrast, we observed a lifespan extension phenotype instead of a lifespan reduction phenotype following upp-1 RNAi (Figure 2B). To confirm this difference, we obtained the upp-1 (jg1, jg2 and jg3) mutant strains from the Jaegal Shim laboratory and verified that the lifespan of the upp-1 mutant worms was shortened (Supplementary Figure 3C). In addition, we constructed another upp-1 RNAi vector (upp-1(##)) and detected lifespan when performing upp-1(##) RNAi. Similar to the result of upp-1 RNAi clones (from the Ahringer libraries), animal exposed reconstructed upp-1(##) RNAi displayed the phenotype of lifespan extension (Supplementary Figure 3A). These two different phenotypes of upp-1 RNAi and upp-1 mutants may be caused by two mechanisms: first, the differences in the food source, HT115 (current study) versus OP50 [47] may contribute to these differences in phenotype [48, 49]; second and most importantly, in the previous study, authors used missense or nonsense upp-1 mutants, which may cause the pyrimidine metabolite to reach an abnormally high level that leads to an abnormal physiological state. In our study, the RNAi treatment might cause a moderate decrease in upp-1 function causing the beneficial accumulation of the intermediates but not at a toxic level that affects the physiological state. This accumulation might activate a negative feedback regulation loop to inhibit the Notch signaling pathway to achieve the increased longevity of C. elegans (Supplementary Figure 5).
Longevity molecules that prevent or attenuate age-related degeneration have long been the ultimate goal of humanity. Many studies have determined that nutrients including, carbohydrates, lipids, proteins, minerals, and vitamins, are essential for the survival of organisms. For example, a previous study reported that increasing glucose intake accelerates aging in yeast and C. elegans [50, 51]. Other researchers found that methionine restriction extends the lifespan of Drosophila by downregulating TOR signaling [52]. As one nutrient class, the effect of nucleotides on aging regulation has been scarcely reported. Here, we found that elevated concentrations of some nucleotide bases (e.g., thymine, orotate, β-aminoisobutyrate, uridine, and cytidine) extend the lifespan of worms. Endogenous metabolites such as pyrimidine intermediates could alter C. elegans lifespan, suggesting that an existing internal mechanism may be accessible to intervention; whether this mechanism can translate into manipulating the aging process in humans requires further investigation. The role of Notch signaling in modulating germline proliferation and longevity in response to changes in nucleotide abundance could prompt future mechanistic studies to discover the connections between nucleotide levels and Notch activity, which will help us not only better understand the detail of lifespan regulation by reproductive signals, but also propose a new avenue for investigating lifespan regulation mechanisms by interfering with endogenous metabolic pathways.
Materials and Methods
Chemicals and strains
The strains used for this publication were obtained from the Caenorhabditis Genetics Center (CGC) (University of Minnesota, USA). Worms were maintained on nematode growth media (NGM) plates at 20 °C with E. coli OP50 bacteria as previously described unless otherwise stated [53]. All of the strains used in this study were: Bristol N2 wild-type, CF1038 daf-16(mu86)I, VC199 sir-2.1(ok434)IV, DA1116 eat-2(ad1116)II, CB1370 daf-2(e1370)III, AA89 daf-12(rh274) X, VC870 nhr-49(gk405) I, CF1903 glp-1(e2144) III and EU1 skn-1(zu67) IV.
All chemicals were purchased from Sigma-Aldrich (Munich, Germany), and resolved in PBS. NGM plates containing compounds were equilibrated overnight before use.
RNA interference experiments
For RNAi gene knockdown experiments, we used E. coli HT115 as the food source for C. elegans as previously described [54]. The clones for dpyd-1, upp-1, dhod-1, and upb-1 were derived from the Ahringer library (Source Bioscience, Nottingham, UK). For upp-1(##) RNAi, the designated fragment was obtained by amplification from genomic DNA with primers (forward, 5′- TTTTGACCCGTTAGTTGTCATGC -3′; reverse, 5′- ATCTCCATCCATACGATTCAAAAGA -3′) and subcloning into the L4440 feeding vector (pPD129.36). Then, the resulting plasmids were transformed into the HT115 (DE3) RNase III-deficient E. coli strain [55]. HT115 bacteria transformed with RNAi vectors (L4440) were grown at 37 °C in LB with 100 μg mL-1 ampicillin. Then, freshly prepared bacteria were spotted on NGM plates with 1 mM isopropyl-B-D-thiogalactoside (IPTG) and 100 μg mL-1 ampicillin. In experiments before treatment, synchronized L1 larvae were transferred to fresh plates with gene-specific RNAi bacteria.
Lifespan assay
All strains were grown on NGM plates for 2-3 generations without starvation. All lifespan assays were performed at 20 °C except CF1903, according to standard protocols and as previously described [53]. In brief, 100 late L4 larvae or young adults were transferred to fresh plates containing 10 μM 5-fluro-2′-deoxyuridine (FUdR, Sigma) and the respective compounds and scored every day. To determine the lifespan of the temperature-sensitive mutant CF1903, L1 worms were incubated at 20 °C for 12 h, then transferred to 25 °C until L4 larvae or young adulthood, and then returned to 20 °C for the remainder of the lifespan [13]. To ensure drug potency, animals were transferred every second day. Experiments were repeated at least twice. The mean, SEM, P value and lifespan value were summarized in Supplementary Table 1 in the Supporting Information.
Movement assay
Movement assays were performed as previously described [22, 53]. Briefly, 100 late L4 larvae or young adults were transferred to fresh plates with or without compound and maintained as described in the lifespan assay. When tapping plates, the worms moving in a continuous, coordinated sinusoidal way were characterized as fast movement; otherwise, the worms were classified as a nonfast movement.
Fertility assay
Single L4 or young adults were transferred to single plates with or without compound and subsequently transferred every 24 h to a fresh plate. The offspring yielded by each worm on each day were allowed to hatch and were counted. For every experiment, more than 30 worms were used, and the experiments were conducted three times.
Quantitative RT-PCR assay
Total RNA was extracted using RNAiso Plus (Takara) based on the phenol-chloroform extraction method. Afterward, the RNA was quantified and converted cDNAs using a high capacity cDNA transcription kit (RK20400, ABclonal) following the manufacturers' protocol. mRNA levels were quantified in a SYBR Green Select Master Mix (RK21203, ABclonal) on a CFX96 real-time system (Biorad). For every experiment, biological triplicates (starting from RNA isolation) and technical triplicates were performed. The results were calculated by the 2-ΔΔCt method, and normalized to the reference genes cdc-42 [56]. P values were calculated using a two-tailed Student’s t test. The primers used in this publication are summarized in Supplementary Table 2 in the Supporting Information.
Germ cell number quantification
Synchronized worms grown on plates with or without treatment were collected 12 h post-L4. Worms from each condition were washed three times with M9 buffer and dissected in M9 buffer with 5 mM imidazole under the dissection light microscope, transferred to a slide and freeze cracked. Gonads were stained using 4,6-diamidino-2-phenylindole (DAPI) as previously described with some modification [57]. Briefly, gonads were fixed in -20 °C MeOH for 10 min, washed twice in PBST (PBS with 0.1% Tween 20), and stained with 0.5 μg/ml DAPI for 10 min. Then, the gonads were washed twice in PBST before imaging. Imaging was performed with a Nikon Ti2-U microscope, and germ cell nuclei were manually counted from the distal tip to the beginning of meiotic entry. At least 20 animals were used for each experiment, and each experiment was repeated at least twice.
Oil red O staining and quantification
ORO staining of fixed worms was conducted as previously described [58]. ORO-stained worms were mounted onto 2% agar pads and imaged at 20× magnification using a Nikon Ti2-U fluorescence microscope. The ORO intensity per worm was measured using Image-Pro-Plus processing software. Mean intensity values in arbitrary units (a.u.) were graphed using GraphPad Prism, and statistical significance was determined using a two-tailed Student’s t test. At least 30 animals were used for each experiment, and each experiment was repeated at least twice.
Green fluorescent protein quantification and visualization
For the quantification of SOD-3 and GST-4, synchronized L1 larvae of CF1553 (muIs84 [Psod-3::GFP, rol-6]) and CL2166 (dvIs19 [(pAF15)gst-4p::GFP::NLS]) were transferred to compounds treated and untreated plates for 55 h. Worms were anesthetized in M9 containing 10 μM levamisole and mounted on 2% agar pads. The GFP fluorescence of worms was directly observed with a Nikon Ti2-U fluorescence microscope. At least 30 animals were used for each experiment. For GFP quantification, images were analyzed by ImageJ.
Determination of pyrimidine intermediates concentrations by HPLC/MS
The strains were cultured for 2-3 generations prior to collection. The samples were prepared as previously described [13]. For sample treated with metabolites, synchronized L1 larvae were grown at the plates with or without metabolites until young adult for harvest. For glp-1 mutants, synchronized L1 larvae were incubated at 20 °C for 12 h, then transferred to 25 °C to eliminate germ cells, until young adult stage for harvest, as well as corresponding control samples were treated via the same procedures. All samples were collected through M9 buffer, and then immediately flash frozen in liquid nitrogen and stored at -80°C until extraction.
Metabolites form C. elegans samples were extracted four times with 800 μL of precooled MeOH / H2O (2:1) using a TissueLyser at 75 Hz for 120 s. All extracts were subjected to centrifugation (12000 rpm for 10 min at 4 °C), supernatant was evaporated to dryness with a vacuo at a low temperature and dissolved in 400 μL MeOH / H2O (2:1) for LC/MS analysis.
Liquid chromatography was performed using a reversed-phase C18 column (Eclipse Plus C18, Agilent, 5μm, 150×2.1 mm diameter column) with a flow rate of 300 μL/min at room temperature, and 3μL of sample was injected. Eluent A was ACN, eluent B was H2O and eluent C was MeOH. The initial eluent consisted of 1% solvent A and 1% solvent C; the percent of buffer A and C were then gradually increased to 10% in 2 min, held there for 4 min, and then returned to the initial condition in 0.1 min, and held there for 10 min, the total run time was 16 min. Analyses were conducted using an Agilent Technologies 1260 (Agilent, Santa Clara, CA) system connected to an AB SCIEX QTRAP 4500 (ESI-MS/MS; Applied Biosystems, Foster City, CA).
Mass spectrometry analyses were conducted in negative ion multiple reaction monitoring (MRM) mode. The MS/MS parameters were optimized by infusion of individual compounds into the MS through a flow injection system.
Acknowledgments
We would like to thank Dr. Xuhui Lai for technical help on our qPCR procedure and Ms. Ruowei Zhong for collecting C. elegans lifespan data. We would like to thank Jaegal Shim for stains upp-1 (jg 1), upp-1 (jg 2) and upp-1 (jg 3); the Caenorhabditis Genetic Center (CGC) for providing the worm strains, which is funded by the NIH Office of Research Infrastructure Programs (P40OD010440).
Conflicts of Interest
There is no conflict of interests exists in the submission of this manuscript, and the manuscript is approved by all authors for publication.
Funding
This work was supported by the National Natural Science Foundation of China (81601299, 81670813), the Program of Introducing Talents of Discipline to Universities (111 Project, No. B16021), Science and Technology Program of Guangzhou (Grant 201607010256) and China Postdoctoral Science Special Fund (2018M633288).
References
- 1. Fontana L, Partridge L. Promoting health and longevity through diet: from model organisms to humans. Cell. 2015; 161:106–18. https://doi.org/10.1016/j.cell.2015.02.020 [PubMed]
- 2. Olshansky SJ. From Lifespan to Healthspan. JAMA. 2018; 320:1323–24. https://doi.org/10.1001/jama.2018.12621 [PubMed]
- 3. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013; 153:1194–217. https://doi.org/10.1016/j.cell.2013.05.039 [PubMed]
- 4. Barzilai N, Crandall JP, Kritchevsky SB, Espeland MA. Metformin as a Tool to Target Aging. Cell Metab. 2016; 23:1060–65. https://doi.org/10.1016/j.cmet.2016.05.011 [PubMed]
- 5. Kennedy BK, Lamming DW. The Mechanistic Target of Rapamycin: The Grand ConducTOR of Metabolism and Aging. Cell Metab. 2016; 23:990–1003. https://doi.org/10.1016/j.cmet.2016.05.009 [PubMed]
- 6. Lucanic M, Held JM, Vantipalli MC, Klang IM, Graham JB, Gibson BW, Lithgow GJ, Gill MS. N-acylethanolamine signalling mediates the effect of diet on lifespan in Caenorhabditis elegans. Nature. 2011; 473:226–29. https://doi.org/10.1038/nature10007 [PubMed]
- 7. Williams DS, Cash A, Hamadani L, Diemer T. Oxaloacetate supplementation increases lifespan in Caenorhabditis elegans through an AMPK/FOXO-dependent pathway. Aging Cell. 2009; 8:765–68. https://doi.org/10.1111/j.1474-9726.2009.00527.x [PubMed]
- 8. Chin RM, Fu X, Pai MY, Vergnes L, Hwang H, Deng G, Diep S, Lomenick B, Meli VS, Monsalve GC, Hu E, Whelan SA, Wang JX, et al. The metabolite α-ketoglutarate extends lifespan by inhibiting ATP synthase and TOR. Nature. 2014; 510:397–401. https://doi.org/10.1038/nature13264 [PubMed]
- 9. Garavito MF, Narváez-Ortiz HY, Zimmermann BH. Pyrimidine Metabolism: Dynamic and Versatile Pathways in Pathogens and Cellular Development. J Genet Genomics. 2015; 42:195–205. https://doi.org/10.1016/j.jgg.2015.04.004 [PubMed]
- 10. Simmonds HA, Duley JA, Fairbanks LD, McBride MB. When to investigate for purine and pyrimidine disorders. Introduction and review of clinical and laboratory indications. J Inherit Metab Dis. 1997; 20:214–26. https://doi.org/10.1023/A:1005308923168 [PubMed]
- 11. Ansoleaga B, Jové M, Schlüter A, Garcia-Esparcia P, Moreno J, Pujol A, Pamplona R, Portero-Otín M, Ferrer I. Deregulation of purine metabolism in Alzheimer’s disease. Neurobiol Aging. 2015; 36:68–80. https://doi.org/10.1016/j.neurobiolaging.2014.08.004 [PubMed]
- 12. Brink TC, Demetrius L, Lehrach H, Adjaye J. Age-related transcriptional changes in gene expression in different organs of mice support the metabolic stability theory of aging. Biogerontology. 2009; 10:549–64. https://doi.org/10.1007/s10522-008-9197-8 [PubMed]
- 13. Wan QL, Shi X, Liu J, Ding AJ, Pu YZ, Li Z, Wu GS, Luo HR. Metabolomic signature associated with reproduction-regulated aging in Caenorhabditis elegans.. Aging (Albany NY). 2017; 9:447–74. https://doi.org/10.18632/aging.101170 [PubMed]
- 14. Lapierre LR, Hansen M. Lessons from C. elegans: signaling pathways for longevity. Trends Endocrinol Metab. 2012; 23:637–44. https://doi.org/10.1016/j.tem.2012.07.007 [PubMed]
- 15. Walther DM, Kasturi P, Zheng M, Pinkert S, Vecchi G, Ciryam P, Morimoto RI, Dobson CM, Vendruscolo M, Mann M, Hartl FU. Widespread Proteome Remodeling and Aggregation in Aging C. elegans. Cell. 2015; 161:919–32. https://doi.org/10.1016/j.cell.2015.03.032 [PubMed]
- 16. Berman JR, Kenyon C. Germ-cell loss extends C. elegans life span through regulation of DAF-16 by kri-1 and lipophilic-hormone signaling. Cell. 2006; 124:1055–68. https://doi.org/10.1016/j.cell.2006.01.039 [PubMed]
- 17. Kenyon CJ. The genetics of ageing. Nature. 2010; 464:504–12. https://doi.org/10.1038/nature08980 [PubMed]
- 18. Tepper RG, Ashraf J, Kaletsky R, Kleemann G, Murphy CT, Bussemaker HJ. PQM-1 complements DAF-16 as a key transcriptional regulator of DAF-2-mediated development and longevity. Cell. 2013; 154:676–90. https://doi.org/10.1016/j.cell.2013.07.006 [PubMed]
- 19. Scott TA, Quintaneiro LM, Norvaisas P, Lui PP, Wilson MP, Leung KY, Herrera-Dominguez L, Sudiwala S, Pessia A, Clayton PT, Bryson K, Velagapudi V, Mills PB, et al. Host-Microbe Co-metabolism Dictates Cancer Drug Efficacy in C. elegans. Cell. 2017; 169:442–456.e418. https://doi.org/10.1016/j.cell.2017.03.040 [PubMed]
- 20. Löffler M, Carrey EA, Zameitat E. Orotic Acid, More Than Just an Intermediate of Pyrimidine de novo Synthesis. J Genet Genomics. 2015; 42:207–19. https://doi.org/10.1016/j.jgg.2015.04.001 [PubMed]
- 21. Martell J, Seo Y, Bak DW, Kingsley SF, Tissenbaum HA, Weerapana E. Global Cysteine-Reactivity Profiling during Impaired Insulin/IGF-1 Signaling in C. elegans Identifies Uncharacterized Mediators of Longevity. Cell Chem Biol. 2016; 23:955–66. https://doi.org/10.1016/j.chembiol.2016.06.015 [PubMed]
- 22. Huang C, Xiong C, Kornfeld K. Measurements of age-related changes of physiological processes that predict lifespan of Caenorhabditis elegans. Proc Natl Acad Sci USA. 2004; 101:8084–89. https://doi.org/10.1073/pnas.0400848101 [PubMed]
- 23. Tullet JM, Hertweck M, An JH, Baker J, Hwang JY, Liu S, Oliveira RP, Baumeister R, Blackwell TK. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. elegans. Cell. 2008; 132:1025–38. https://doi.org/10.1016/j.cell.2008.01.030 [PubMed]
- 24. Ogg S, Paradis S, Gottlieb S, Patterson GI, Lee L, Tissenbaum HA, Ruvkun G. The Fork head transcription factor DAF-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature. 1997; 389:994–99. https://doi.org/10.1038/40194 [PubMed]
- 25. Van Gilst MR, Hadjivassiliou H, Yamamoto KR. A Caenorhabditis elegans nutrient response system partially dependent on nuclear receptor NHR-49. Proc Natl Acad Sci USA. 2005; 102:13496–501. https://doi.org/10.1073/pnas.0506234102 [PubMed]
- 26. Motola DL, Cummins CL, Rottiers V, Sharma KK, Li T, Li Y, Suino-Powell K, Xu HE, Auchus RJ, Antebi A, Mangelsdorf DJ. Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell. 2006; 124:1209–23. https://doi.org/10.1016/j.cell.2006.01.037 [PubMed]
- 27. Murphy CT, McCarroll SA, Bargmann CI, Fraser A, Kamath RS, Ahringer J, Li H, Kenyon C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature. 2003; 424:277–83. https://doi.org/10.1038/nature01789 [PubMed]
- 28. McCormick M, Chen K, Ramaswamy P, Kenyon C. New genes that extend Caenorhabditis elegans’ lifespan in response to reproductive signals. Aging Cell. 2012; 11:192–202. https://doi.org/10.1111/j.1474-9726.2011.00768.x [PubMed]
- 29. Folick A, Oakley HD, Yu Y, Armstrong EH, Kumari M, Sanor L, Moore DD, Ortlund EA, Zechner R, Wang MC. Aging. Lysosomal signaling molecules regulate longevity in Caenorhabditis elegans. Science. 2015; 347:83–86. https://doi.org/10.1126/science.1258857 [PubMed]
- 30. Chi C, Ronai D, Than MT, Walker CJ, Sewell AK, Han M. Nucleotide levels regulate germline proliferation through modulating GLP-1/Notch signaling in C. elegans. Genes Dev. 2016; 30:307–20. https://doi.org/10.1101/gad.275107.115 [PubMed]
- 31. Berry LW, Westlund B, Schedl T. Germ-line tumor formation caused by activation of glp-1, a Caenorhabditis elegans member of the Notch family of receptors. Development. 1997; 124:925–36. [PubMed]
- 32. Arantes-Oliveira N, Apfeld J, Dillin A, Kenyon C. Regulation of life-span by germ-line stem cells in Caenorhabditis elegans. Science. 2002; 295:502–05. https://doi.org/10.1126/science.1065768 [PubMed]
- 33. Ratnappan R, Amrit FR, Chen SW, Gill H, Holden K, Ward J, Yamamoto KR, Olsen CP, Ghazi A. Germline signals deploy NHR-49 to modulate fatty-acid β-oxidation and desaturation in somatic tissues of C. elegans. PLoS Genet. 2014; 10:e1004829. https://doi.org/10.1371/journal.pgen.1004829 [PubMed]
- 34. Angelo G, Van Gilst MR. Starvation protects germline stem cells and extends reproductive longevity in C. elegans. Science. 2009; 326:954–58. https://doi.org/10.1126/science.1178343 [PubMed]
- 35. Wang MC, O’Rourke EJ, Ruvkun G. Fat metabolism links germline stem cells and longevity in C. elegans. Science. 2008; 322:957–60. https://doi.org/10.1126/science.1162011 [PubMed]
- 36. Berdichevsky A, Viswanathan M, Horvitz HR, Guarente L. C. elegans SIR-2.1 interacts with 14-3-3 proteins to activate DAF-16 and extend life span. Cell. 2006; 125:1165–77. https://doi.org/10.1016/j.cell.2006.04.036 [PubMed]
- 37. Hansen M, Chandra A, Mitic LL, Onken B, Driscoll M, Kenyon C. A role for autophagy in the extension of lifespan by dietary restriction in C. elegans. PLoS Genet. 2008; 4:e24. https://doi.org/10.1371/journal.pgen.0040024 [PubMed]
- 38. Heestand BN, Shen Y, Liu W, Magner DB, Storm N, Meharg C, Habermann B, Antebi A. Dietary restriction induced longevity is mediated by nuclear receptor NHR-62 in Caenorhabditis elegans. PLoS Genet. 2013; 9:e1003651. https://doi.org/10.1371/journal.pgen.1003651 [PubMed]
- 39. McKay JP, Raizen DM, Gottschalk A, Schafer WR, Avery L. eat-2 and eat-18 are required for nicotinic neurotransmission in the Caenorhabditis elegans pharynx. Genetics. 2004; 166:161–69. https://doi.org/10.1534/genetics.166.1.161 [PubMed]
- 40. Nyhan WL. Disorders of purine and pyrimidine metabolism. Mol Genet Metab. 2005; 86:25–33. https://doi.org/10.1016/j.ymgme.2005.07.027 [PubMed]
- 41. Maring JG, Groen HJ, Wachters FM, Uges DR, de Vries EG. Genetic factors influencing pyrimidine-antagonist chemotherapy. Pharmacogenomics J. 2005; 5:226–43. https://doi.org/10.1038/sj.tpj.6500320 [PubMed]
- 42. Chi C, Han M. Notch signaling protects animals from nucleotide deficiency. Cell Cycle. 2016; 15:1941–42. https://doi.org/10.1080/15384101.2016.1181878 [PubMed]
- 43. Häberle J, Rubio V. Disorders of the urea cycle and related enzymes. Inborn Metabolic Diseases. Springer; 2016. pp. 295–308. https://doi.org/10.1007/978-3-662-49771-5_19
- 44. Simmonds HA. Enzymes of nucleotide biosynthesis: differences between intact and lysed cells as well as between species and tissues can be important. Biochem Soc Trans. 1995; 23:877–9. https://doi.org/10.1042/bst0230877 [PubMed]
- 45. Page T, Yu A, Fontanesi J, Nyhan WL. Developmental disorder associated with increased cellular nucleotidase activity. Proc Natl Acad Sci USA. 1997; 94:11601–06. https://doi.org/10.1073/pnas.94.21.11601 [PubMed]
- 46. Yan R, Wan L, Pizzorno G, Cao D. Uridine phosphorylase in breast cancer: a new prognostic factor? Front Biosci. 2006; 11:2759–66. https://doi.org/10.2741/2005 [PubMed]
- 47. Kim S, Park DH, Kim TH, Hwang M, Shim J. Functional analysis of pyrimidine biosynthesis enzymes using the anticancer drug 5-fluorouracil in Caenorhabditis elegans. FEBS J. 2009; 276:4715–26. https://doi.org/10.1111/j.1742-4658.2009.07168.x [PubMed]
- 48. Pang S, Curran SP. Adaptive capacity to bacterial diet modulates aging in C. elegans. Cell Metab. 2014; 19:221–31. https://doi.org/10.1016/j.cmet.2013.12.005 [PubMed]
- 49. Xiao R, Chun L, Ronan EA, Friedman DI, Liu J, Xu XZ. RNAi Interrogation of Dietary Modulation of Development, Metabolism, Behavior, and Aging in C. elegans. Cell Rep. 2015; 11:1123–33. https://doi.org/10.1016/j.celrep.2015.04.024 [PubMed]
- 50. Lee SJ, Murphy CT, Kenyon C. Glucose shortens the life span of C. elegans by downregulating DAF-16/FOXO activity and aquaporin gene expression. Cell Metab. 2009; 10:379–91. https://doi.org/10.1016/j.cmet.2009.10.003 [PubMed]
- 51. Gusarov I, Pani B, Gautier L, Smolentseva O, Eremina S, Shamovsky I, Katkova-Zhukotskaya O, Mironov A, Nudler E. Glycogen controls Caenorhabditis elegans lifespan and resistance to oxidative stress. Nat Commun. 2017; 8:15868. https://doi.org/10.1038/ncomms15868 [PubMed]
- 52. Lee BC, Kaya A, Ma S, Kim G, Gerashchenko MV, Yim SH, Hu Z, Harshman LG, Gladyshev VN. Methionine restriction extends lifespan of Drosophila melanogaster under conditions of low amino-acid status. Nat Commun. 2014; 5:3592. https://doi.org/10.1038/ncomms4592 [PubMed]
- 53. Wan QL, Zheng SQ, Wu GS, Luo HR. Aspirin extends the lifespan of Caenorhabditis elegans via AMPK and DAF-16/FOXO in dietary restriction pathway. Exp Gerontol. 2013; 48:499–506. https://doi.org/10.1016/j.exger.2013.02.020 [PubMed]
- 54. Mansfeld J, Urban N, Priebe S, Groth M, Frahm C, Hartmann N, Gebauer J, Ravichandran M, Dommaschk A, Schmeisser S, Kuhlow D, Monajembashi S, Bremer-Streck S, et al. Branched-chain amino acid catabolism is a conserved regulator of physiological ageing. Nat Commun. 2015; 6:10043. https://doi.org/10.1038/ncomms10043 [PubMed]
- 55. Timmons L, Fire A. Specific interference by ingested dsRNA. Nature. 1998; 395:854. https://doi.org/10.1038/27579 [PubMed]
- 56. Hoogewijs D, Houthoofd K, Matthijssens F, Vandesompele J, Vanfleteren JR. Selection and validation of a set of reliable reference genes for quantitative sod gene expression analysis in C. elegans. BMC Mol Biol. 2008; 9:9. https://doi.org/10.1186/1471-2199-9-9 [PubMed]
- 57. Käser-Pébernard S, Müller F, Wicky C. LET-418/Mi2 and SPR-5/LSD1 cooperatively prevent somatic reprogramming of C. elegans germline stem cells. Stem Cell Reports. 2014; 2:547–59. https://doi.org/10.1016/j.stemcr.2014.02.007 [PubMed]
- 58. O’Rourke EJ, Soukas AA, Carr CE, Ruvkun G. C. elegans major fats are stored in vesicles distinct from lysosome-related organelles. Cell Metab. 2009; 10:430–35. https://doi.org/10.1016/j.cmet.2009.10.002 [PubMed]