



Introduction
Aging remains associated with a variety of diseases, including infertility, degenerative diseases, and cancer [1]. Female ovarian reserve has been reported to decline progressively with advancing age [2]. Women in the elderly age group have a decreased follicle pool of less than 1,000 and show irregular ovulatory cyclic changes, the first clinical signs of ovarian aging [3, 4]. Ovarian aging results in a poor response to ovarian stimulation leading to reduced number and quality of remaining oocytes in women [5, 6]. In particular, older women (> 40 years), are subjected to increased risks of chromosomal abnormalities in oocytes and adverse pregnancy outcomes [7, 8]. The most commonly faced challenge has been the effective management and improvement of age-related decline in female reproductive potential [9].
Recent research has focused on developing effective fertility preservation strategies, such as ovarian tissue and oocyte cryopreservation [10]. However, such attempts are likely to be effective only in women younger than 35 years and with good ovarian reserve. Because older women with poor ovarian responses (POR) have a declining oocyte yield in in vitro fertilization, high-dose follicle-stimulating hormone (FSH) protocols are unsuccessful [11]. Thus, differential pharmacological strategies are required for women to improve the ovarian reserve.
Alternative options in integrative medicine treatment using natural herbs are considered effective for elderly women with POR. Herbal combinations have been used to manage infertility symptoms in human and rodent models [12–14]. Samul-tang (SM) is a well-known mixed herbal medicine comprising equal proportions of Paeonia lactiflora, Liqusticum striatum, Rehmannia glutinosa, and Angelica gigas. Pharmacological effects of SM include anti-inflammatory, antioxidative, anti-stress, and anti-cancer effects [15]. The herb is commonly prescribed to women with gynecological disorders, such as irregular menstruation and postmenopausal syndrome, often clinically observed in older women with POR [16]. Anti-aging effects of three compounds, including P. lactiflora, R. glutinosa, and A. gigas, have been reported using human cells and animal models [17–19]. Although herbal medicines are traditionally known to improve fertility-related parameters, scientific evidence supporting their use for age-related infertility is limited.
In this study, the influence of SM administration on ovarian reserve for four weeks in old-aged mice was investigated. Further, the compensatory mechanisms of SM in age-related functional decline in ovary were uncovered using transcriptomic analysis.
Results
SM improved age-related decline of serum AMH and FSH level
Body weight changes in mice of each group were measured for four weeks since SM administration (Figure 1A). SM had no effect on the body weight (Figure 1B). However, decreased ovary weight associated with aging was regained by SM administration for four weeks (Figure 1C).
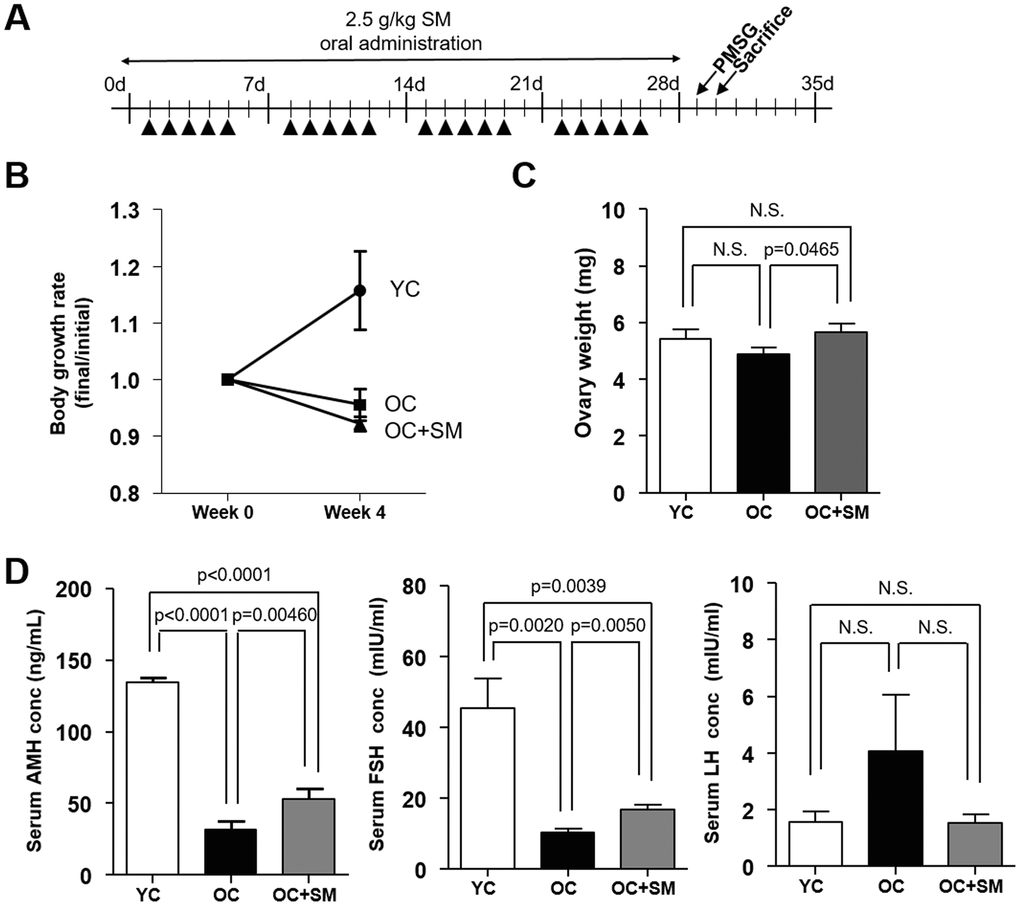
Figure 1. Body weight changes and serum hormone levels in mice after Samul-tang (SM) administration. (A) Eight-week-old mice were orally administered distilled water (n=6, YC group). Forty-week-old mice were orally administered distilled water (n=6, OC group) or 2.5 g/kg of SM (n= 7, OC+SM group) five times a week for four weeks. Post SM administration, the mice were weighed and hormonal assessment was performed. (B) Body weight changes. (C) Ovary weight. (D) Serum levels of anti-Müllerian hormone (AMH), follicle-stimulating hormone (FSH), and luteinizing hormone(LH). Data are presented as mean ± standard error of the mean. Statistical analysis was performed using the Student’s t-test.
Next, we measured the serum levels of anti-Müllerian hormone (AMH), FSH, and luteinizing hormone (LH) in each mouse group. OC mice (40-week-old mice orally administered distilled water) exhibited low AMH levels resulting in a decreased ovarian reserve (Figure 1D). SM administration in OC+SM mice significantly enhanced the serum AMH levels. Although serum LH levels did not change, FSH levels increased significantly in OC+SM mice compared to those in OC mice (Figure 1D). It is likely that the production of more FSH directly stimulates the granulosa cells for follicular growth and development [20].
SM prevented aged-related ovarian follicle loss
To investigate the protective effects of SM on ovarian follicle growth under aging conditions, a histological analysis of the ovaries excised from the mice after SM administration was performed (Figure 2A). Histological changes in YC (8-week-old mice orally administered distilled water), OC, and OC+SM mice were observed in ovarian tissues (Figure 2B). Compared to the YC mice, aged mice showed an overall reduction in ovarian follicles. OC+SM mice showed higher number of follicles at all stages than the OC mice (Figure 2C). These results suggest that SM administration prevents depletion of the primordial follicle pool.
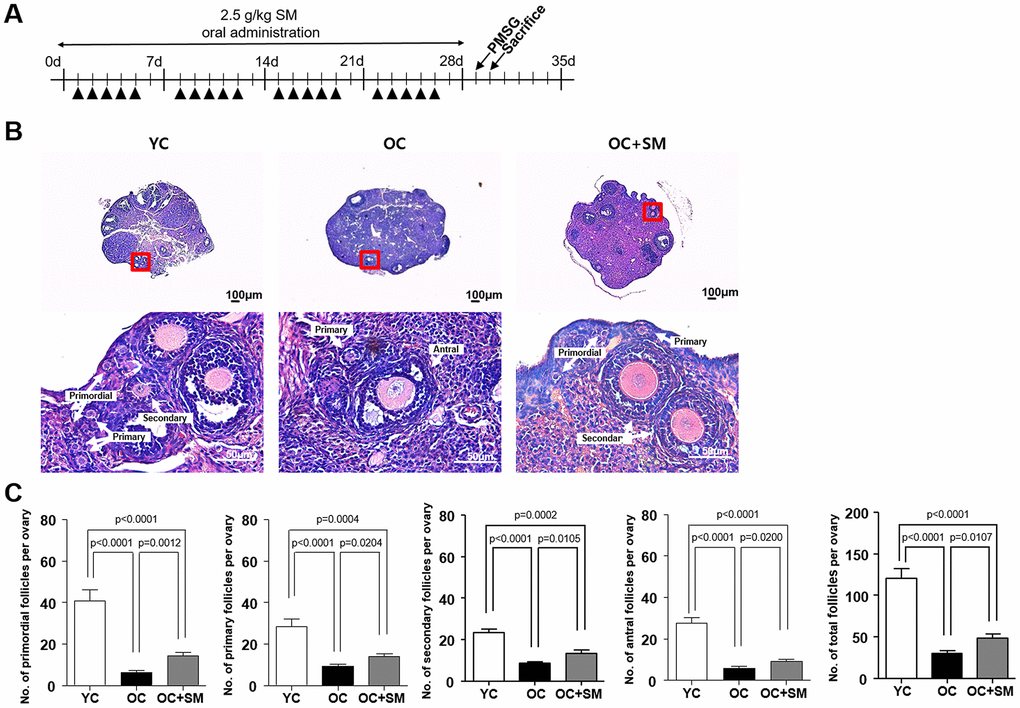
Figure 2. Histological analysis of ovarian follicles in mice after Samul-tang (SM) administration. (A) Eight-week-old mice were orally administered distilled water (n = 6, YC group). Forty-week-old mice were orally administered distilled water (n = 6, OC group) or 2.5 g/kg of SM (n = 7, OC+SM group) five times a week for four weeks. (B) Post SM administration, both mouse ovaries were assessed histologically. (C) Number of ovarian follicles in different stages and the total number of ovarian follicles. Data are presented as mean ± standard error of the mean. Statistical analysis was performed using the Student’s t-test.
SM ameliorated aged-related impairment of oocyte quality
To investigate the protective effects of SM on oogenesis under aging conditions, mice were hormonally superovulated to collect the oocytes (Figure 3A). Thereafter, the quantity and quality of oocytes were assessed (Figure 3B). Number of total oocytes retrieved from the aged mice was significantly lower than that retrieved from the YC mice (P< 0.05; Figure 3C). However, the number of mature metaphase II (MII) oocytes, with normal chromosomes and well-organized spindle alignments, was significantly higher in OC+SM mice than in OC mice (Figure 3D, 3E). These results indicate that SM administration could improve the quality of oocytes in aged mice.
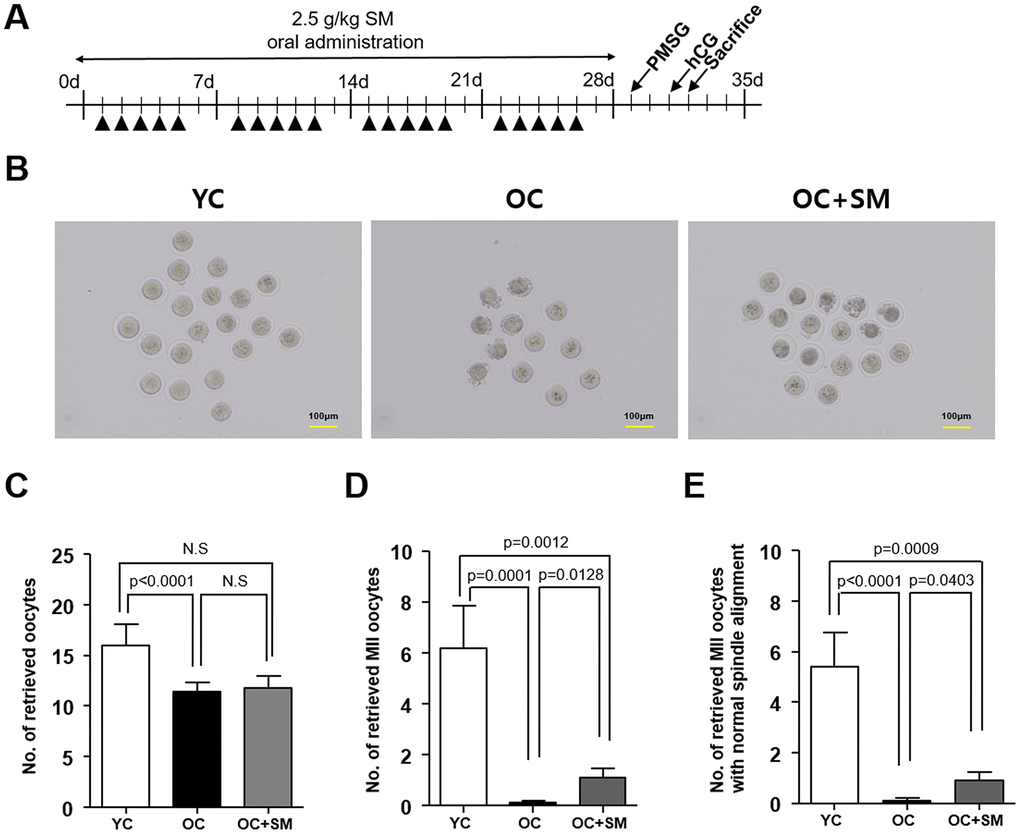
Figure 3. Quality and quantity of mouse oocytes retrieved after Samul-tang (SM) administration. (A) Eight-week-old mice were orally administered distilled water (n = 6, YC group). Forty-week-old mice were orally administered distilled water (n = 10, OC group) or 2.5 g/kg of SM (n = 10, OC+SM group) five times a week for four weeks. Post SM administration, the mice were superovulated via hormonal stimulation. (B) Oocytes retrieved from the YC, OC, and OC+SM mice at 18 h after hCG injection. Number of retrieved oocytes (C), mature metaphase II (MII) oocytes (D), and MII oocytes with normal chromosomal and spindle alignment (E), retrieved from the three different groups of mice. Data are presented as mean ± standard error of the mean. Statistical analysis was performed using the Student’s t-test.
SM reversed aged-induced changes in mRNA expression in mouse ovaries
QuantSeq 3’ messenger RNA (mRNA) sequencing was performed to compare the mRNA expression patterns in ovulated ovaries obtained from YC, OC, and OC+SM mice (Figure 4A). Hierarchical clustering analysis revealed 2,389 marked differentially expressed genes (DEGs) in the three mice groups, with fold changes >1.5 (P< 0.05; Figure 4B). We specifically focused on 393 DEGs in the OC+SM vs. OC group (Figure 4C; blue circle). Bioinformatic analysis was performed for 122 upregulated genes (31.0%) and 271 downregulated genes (69.0%) (Supplementary Tables 1, 2). Kyoto Encyclopedia of Genes and Genomes pathway analysis of OC and OC+SM mice datasets revealed that the DEGs were involved in the RAS signaling pathway (Table 1). Expression of nine genes associated with the RAS signaling pathway was analyzed. The genes include fibroblast growth factor receptor 2 (Fgfr2), guanine nucleotide binding protein gamma 8 (Gng8), RAS protein activator like 1 (Rasal1), RAS protein-specific guanine nucleotide-releasing factor 1 and 2 (Rasgrf1 and Rasgrf2), neurofibromatosis 1 (Nf1), fibroblast growth factor 13 (Fgf13), Src homology 2 domain-containing transforming protein C3 (Shc3), and epidermal growth factor (Egf). Fgfr2, Rasgrf2, and Egf were downregulated in OC mice; however, their expression levels were restored with SM administration (Figure 5A). Likewise, SM administration also restored the expression levels of Shc3 and Rasal1, which were initially upregulated in OC mice (Figure 5B). Expression of Nf1 and Fgf13 in the ovaries of OC+SM mice decreased significantly compared to that in the ovaries of YC and OC mice (Figure 5C). Gng8 and Rasgrf1 expressions increased significantly in OC+SM mice than in the ovaries of OC mice (Figure 5C). These genes were validated by quantitative polymerase chain reaction (qPCR) using the fluorescent probe-based TaqMan assay (Figure 6). Different expression patterns of Gng8 and Rasgrf1 were observed in sequencing data. Given the filtering of transcriptomics technologies for quality measures, including fold-change >1.5 and P < 0.05, this study obtained varying results from sequencing and qPCR analysis [21]. These findings suggest that SM administration ameliorates the age-related deterioration of ovarian function via genetic regulation.
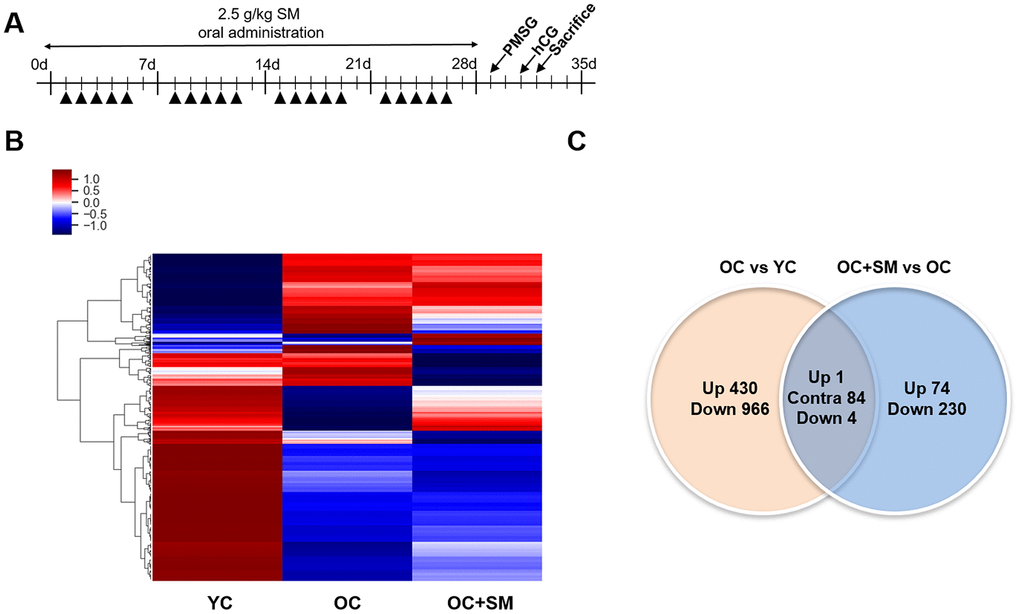
Figure 4. Hierarchical clustering and analysis of differentially expressed messenger RNAs (mRNAs). (A) QuantSeq 3’ mRNA analysis was performed to compare the gene expression in ovulated ovaries of YC (n = 6), OC (n = 6), and OC+SM (n = 6) mice. (B) Hierarchical clustering among the mRNA expression profiles showing 2,389 differentially expressed mRNAs in the three groups, with a fold-change >1.5 and P< 0.05. (C) Venn diagram presenting the numbers of differentially expressed mRNAs between OC vs. YC and OC+SM vs. OC pairs. YC: 8-week-old control mice; OC: 40-week-old mice; OC+SM: 40-week-old mice orally administered Samul-tang. Up, upregulated genes between compared sets; Contra, contraregulated genes between compared sets; Down, downregulated genes between compared sets.
Table 1. Functional annotation of differentially expressed genes.
| Term | P-value | Genes | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mmu05034: Alcoholism | 5.58E-04 | Gng8, Hist4h4, Hist1h4a, Npy, Hist1h2bf, Hist2h2ac, Creb3l2, Hist1h4d, shc3, Hist1h2bq | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mmu04014: Ras signaling pathway | 0.00051931 | Fgfr2, Gng8, Rasal1, Rasgrf1, Rasgrf2, Nf1, Fgf13, Shc3, Egf | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mmu04911: Insulin secretion | 0.0179786 | Gcg, Slc2a2, Creb3l2, Cacna1f, Pclo | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mmu05322: Systemic lupus erythematosus | 0.0286335 | Hist4h4, Hist1h4a, Hist1h2bf, Hist2h2ac, Hist1h4d, Hist1h2bq | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kyoto Encyclopedia of Genes and Genomes analysis was performed to identify the potential signaling pathway of the differentially expressed genes. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
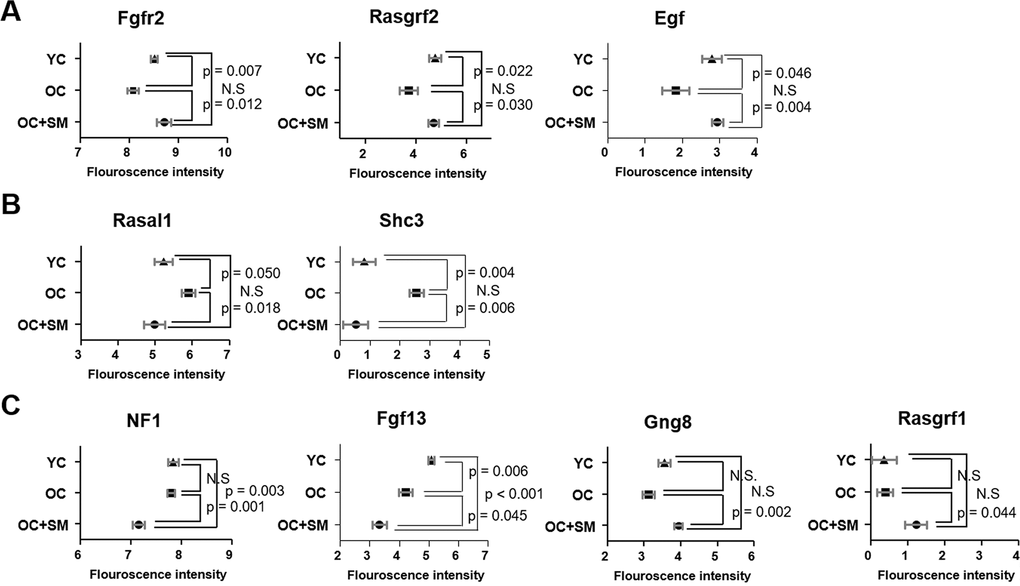
Figure 5. Differentially expressed mRNAs involved in RAS signaling pathway. Fluorescence intensities indicating the expression levels of Fgfr2, Rasgrf1, Egf, Rasal1, Sch3, Nf1, Fgf13, Gng8, and Rasgrf2 in the ovaries of YC, OC, and OC+SM mice. (A) Aging-induced downregulated expression of Fgfr2, Rasgrf2 and Egf were restored with SM administration. (B) Aging-induced upregulated expression of Rasal1 and Sch3 were restored with SM administration. (C) Expression of Nf1, Fgf13, Gng8, and Rasgrf1 were changed by SM administration in OC mice. Data are presented as mean ± standard error of the mean. Statistical analysis was performed using the Student’s t-test. YC: 8-week-old control mice; OC: 40-week-old mice; OC+SM: 40-week-old mice orally administered Samul-tang.
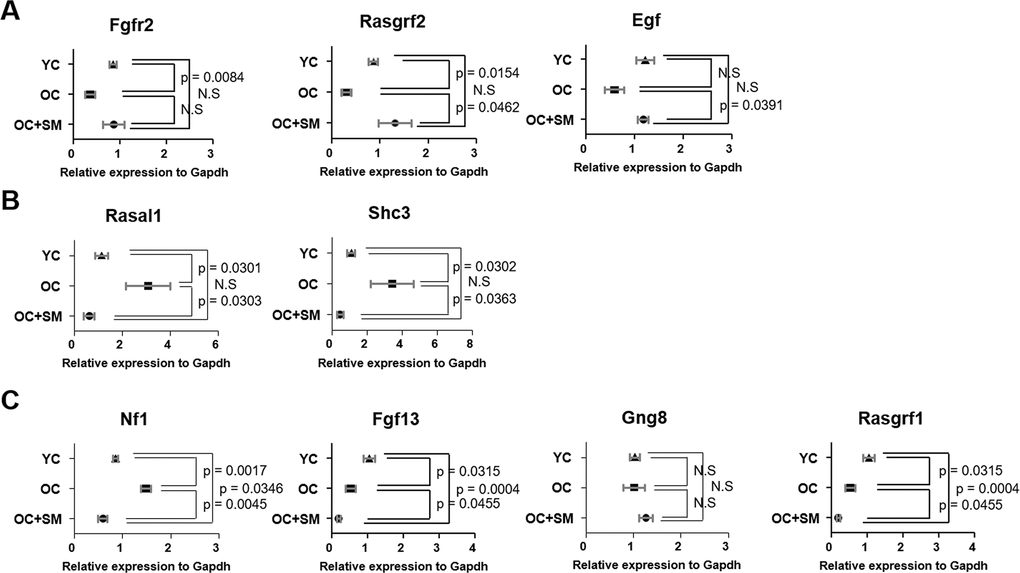
Figure 6. Validation of expression of differentially expressed mRNAs involved in ovarian function. Quantitative polymerase chain reaction was performed to validate the expression of Fgfr2, Rasgrf1, Egf, Rasal1, Sch3, Nf1, Fgf13, Gng8, and Rasgrf2 in the ovaries of YC, OC and OC+SM mice. (A) Aging-induced downregulated expression of Fgfr2, Rasgrf2 and Egf were restored with SM administration. (B) Aging-induced upregulated expression of Rasal1 and Sch3 were restored with SM administration. (C) Expression of Nf1, Fgf13, Gng8, and Rasgrf1 were changed by SM administration in OC mice. Data are presented as mean ± standard error of the mean. Statistical analysis was performed using the Student’s t-test. YC: 8-week-old control mice; OC: 40-week-old mice; OC+SM: 40-week-old mice orally administered Samul-tang.
SM improved aged-induced decline in blastocyst developmental competence
To investigate whether SM enhances the developmental competence under aging conditions, superovulated mice were mated with fertile males. At 1.5 days post coitum (dpc), 2-cell embryos were flushed from the oviduct and cultured to obtain blastocysts for three days. At 4.5 dpc, blastomeres and DNA fragmentation in blastocysts were stained to assess developmental competence (Figure 7A).
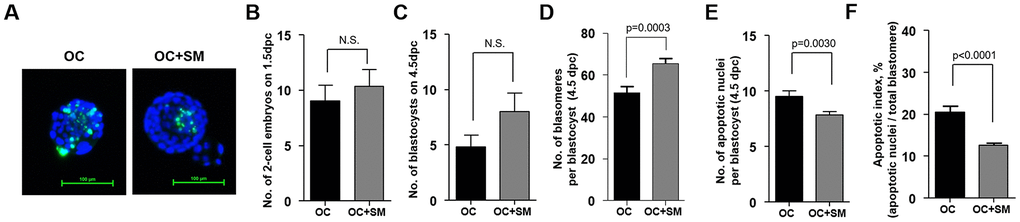
Figure 7. Quality and quantity of mouse blastocysts retrieved after Samul-tang (SM) administration. Forty-week-old mice were orally administered distilled water (n = 13, OC group) or 2.5 g/kg of SM (n =13, OC+SM group) five times a week for four weeks. Post SM administration, the mice were superovulated via hormonal stimulation and mated with fertile males. At 1.5 days post coitum (dpc), 2-cell embryos were collected and cultured up to blastocyst stage for three days. (A) Blastocyst quality was evaluated by assessing the stained nuclei and DNA fragmentation in blastocysts from each mice group. Number of collected 2-cell embryos at 1.5 dpc (B) and cultured blastocysts at 4.5 dpc (C). (D–F) Blastocyst developmental competence was assessed from the number of blastomeres and apoptotic nuclei, and apoptotic index. Data are presented as mean ± standard error of the mean. Statistical analysis was performed using the Student’s t-test.
No significant difference in the number of 2-cell embryos and developed blastocysts was observed between 1.5 to 4.5 dpc (Figure 7B, 7C). However, the number of blastomeres per blastocyst increased significantly. Number of apoptotic cells was lower in OC+SM mice than in OC mice (Figure 7D, 7E). Apoptotic index decreased in blastocysts obtained from OC+SM mice compared to OC mice (Figure 7F). These results indicate that blastocysts with elevated total cell number and decreased apoptotic index could affect the implantation potential in vivo and pregnancy outcomes.
SM improved implantation potential in aged mice
To investigate whether SM enhances the implantation potential in vivo under aging conditions, superovulated mice were mated with fertile males. Pregnancy rate, number and weight of litter were assessed at 9.5 dpc (Figure 8A).
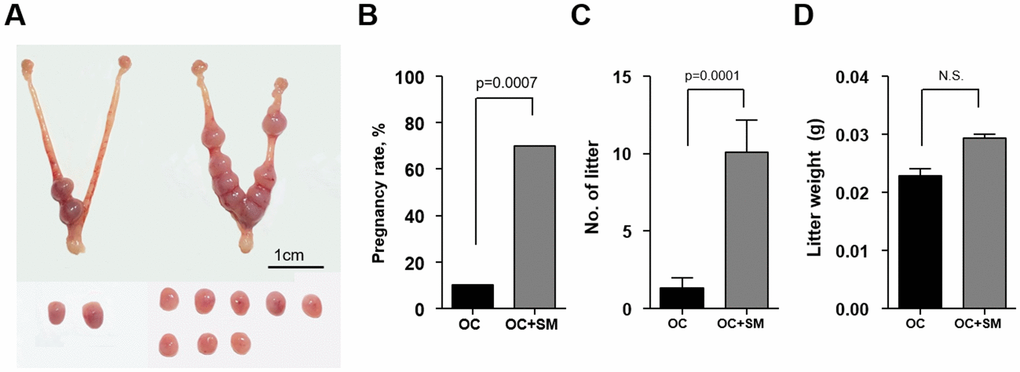
Figure 8. In vivo implantation potential after Samul-tang (SM) administration. Forty-week-old mice were orally administered distilled water (n = 20, OC group) or 2.5 g/kg of SM (n = 10, OC+SM group) five times a week for four weeks. Post SM administration, the mice were superovulated via hormonal stimulation and mated with fertile males. (A) At 9.5 days post coitum (dpc), the uterus from mice of both the groups was collected to assess the implantation potential. (B) Pregnancy rates. (C) Number of litters. (D) Weight of litters. Data are presented as mean ± standard error of the mean. Statistical analysis was performed using the Student’s t-test. The significance of difference in pregnancy rates was determined using Fisher’s exact test.
The pregnancy rate was significantly higher in OC+SM mice than in OC mice (Figure 8B). Furthermore, SM administration enhanced the number of litters having weight similar to OC mice weight (Figure 8C, 8D).
Discussion
Age-related fertility is characterized by gradual decline in ovarian follicle quantity and oocyte quality since birth [22]. Decrease in fertility enhances the risk of reproductive failure [23]. Pharmacological research on anti-ovarian aging agents for women who suffer from POR or premature ovarian failure has gained significant momentum [24]. Thus, new strategies or approaches to prevent ovarian aging are urgently needed.
Progressive decline in tissue homeostasis and biological function with aging contributes to increased risk of miscarriage, degenerative diseases, and death [25]. Some of the well-known causes of aging include oxidative damage caused by generation of free radicals and reactive oxygen species (ROS), and genetic instability-driven genome damage [26]. Oxidative deterioration of DNA, proteins, and lipids by ROS and free radical overproduction has been reported in aged ovaries [27]. Genetic defects in DNA repair systems are considered responsible for the senescent phenotype causing aging syndromes, such as premature aging [28, 29]. SM comprises numerous active components, including gallic acid, paeoniflorin, and ferulic acid. These compounds contain several hydroxyl groups that scavenge the overproduced free radicals [30, 31].
RAS signaling modulates the balance between ROS and antioxidants to prevent cellular senescence or apoptosis [32, 33]. Balanced GDP/GTP cycling maintains the RAS-mediated signaling, which plays a critical role in controlling the normal cellular proliferation and survival. RAS also plays a critical role in the events associated with ovulation and luteinization in preovulatory follicles [34]. Its activity is regulated by multiple guanine nucleotide exchange factors, such as RAS-GRF2 and GTPase-activating proteins, such as Nf1 and Rasal1 [35]. The qPCR results revealed that downregulated Rasgrf2 and upregulated Rasal1 and Shc3 inactivate the RAS-mediated signaling cascades in ovarian follicle development and oocyte maturation in aged ovaries. Interestingly, the altered expression of these genes was restored with SM administration in aged mouse ovaries. EGF also acts through the RAS/ERK pathway to regulate protein homeostasis by promoting the expression of antioxidant genes [36]. EGF signaling is not only essential for normal ovarian steroid oogenesis and oocyte maturation but is also required to induce cumulus cell expansion in vitro during the ovulatory process [37]. Egf expression in aged ovaries receiving SM administration was significantly upregulated compared to that in untreated ovaries. SM likely prevents cell senescence and enhances ovarian reserve by activating Ras in aged women.
Active RAS-mitogen-activated protein kinase pathway can largely regulate FGF signaling, which is known to be associated with several developmental processes, including cellular proliferation, differentiation, angiogenesis, and migration [38]. Fgf13, one of the genes whose expression is decreased by SM, plays an important role in the reconstruction and degradation of the extracellular matrix in infertile aged women [39]. In addition, bidirectional dialog between Fgf2 in oocytes and several Fgfr in granulosa cells is essential for early ovarian folliculogenesis and granulosa cell growth [40]. Restored Fgfr2 expression by SM could pose as an evidence for potential oocyte developmental competence. Functional assays revealed that loss of endothelial Fgfr1 and Fgfr2 results in impaired neovascularization in adult mice [41]. Aged patients exhibit decreased ovarian stromal blood flow owing to a marked reduction in the number and caliber of blood vessels with changes in endothelial cells [42]. Insufficient supply of growth factors owing to decreased blood flow causes follicular arrest and ovarian fibrosis in aged women with POR [43, 44]. SM-induced changes in FGF signaling could drive tissue repair in aged ovaries pre- or post-ovulation [45, 46].
Interestingly, the pregnancy rates were higher in aged mice receiving SM administration. Successful implantation requires good quality of implanting embryo and appropriate structural and functional remodeling of the endometrium [47]. Although SM administration did not increase the number of 2-cell embryos and blastocysts, it enhanced the scope to implant competent blastocysts with more cell divisions.
This study aimed to prevent age-specific decline in functional ovarian reserve using an aged mouse model. However, functional analysis and clinical studies are still warranted to develop strategies or approaches that could improve conditions or delay ovarian aging. Identification of the potential regulatory genes highlighted the SM-induced epigenetic regulation in age-related cognitive decline in ovarian reserve and implantation potential. SM restores the homeostatic balance to revitalize ovarian function under aging condition through the RAS signaling pathway. In conclusion, SM could be helpful in increasing the oocyte yield in aged women by potentially improving the age-related cognitive decline in ovarian reserve.
Materials and Methods
Mice
All experiments and analyses were conducted in accordance with the relevant guidelines and regulations. Experimental protocols concerning animals were approved by the Institutional Animal Care and Use Committee of the Korea Institute of Oriental Medicine, Daejeon, Korea (approval number 19-061). Female BALB/c mice aged 8 and 40 weeks (Central Lab Animal Inc., Seoul, Korea) were housed under specific pathogen-free conditions. To investigate the effect of SM (Hanpoong, Iksan, Korea), the old mice were orally administered either distilled water (OC group) or 2.5 g/kg of SM (OC+SM group) five times a week for four weeks.
Mice from the OC and OC+SM groups were separated within the same cage to synchronize their hormonal cycles [48]. All mice were sacrificed after completion of SM administration. Ovaries were removed, weighed, and immediately placed in 4% paraformaldehyde (Biosesang, Seongnam, Korea) or liquid nitrogen for histological or RNA sequencing, respectively.
Enzyme-linked immunosorbent assay (ELISA) for hormonal assessment
Post SM administration, blood samples were collected from the mice and sera were separated and stored at -80 ° C until analysis. The serum concentrations of AMH, FSH, and LH were measured using hormone-specific ELISA kits from Ansh Lab (Webster, TX, USA), Cusabio Biotech Co. (Wuhan, China), and Endocrine technologies (Newark, CA, USA), respectively, according to the standard protocols and manufacturers’ instructions. For AMH, the inter-assay coefficients of variation (CV) was <10%, with a sensitivity of 0.06 ng/mL. For FSH, both intra- and inter-assay CVs were <15% with a sensitivity of 2.5 mIU/mL. For LH, the intra- and inter-assay CVs were 7% and 15%, respectively, and the functional sensitivity was 5.2 mIU/mL.
Histological assessment of ovarian follicles
Post SM administration, ovaries from YC, OC and OC+SM groups were serially sectioned to obtain 5-μm-thick tissue sections. These sections were then subjected to hematoxylin and eosin staining. Primordial, primary, secondary, and preovulatory follicles, with visible oocytes, were counted in every tenth stained section to avoid repeated counting of the same follicle. The follicular stages were classified as previously described [49]: primordial follicles, with a single flat layer of granulosa cells surrounding the oocyte; primary follicles, with a single cuboidal granulosa cell layer; secondary follicles, with at least two granulosa cell layers and a theca layer; and preovulatory follicles, with a complete antrum and theca layer.
Assessment of oocyte quantity and quality
Post SM administration, the mice were superovulated with an intraperitoneal injection of 5 IU of pregnant mare serum gonadotropin (PMSG; Prospec, Rehovot, Israel). Another injection of 5 IU of human chorionic gonadotropin (hCG; Prospec) was administered 48 h later. Oocytes were collected 18 h post-hCG injection in preincubated human tubal fluid medium (Irvine Scientific, CA, USA). Oocytes were fixed with 4% paraformaldehyde (Biosesang), permeabilized with 0.5% Triton X-100 (Sigma–Aldrich, St. Louis, MO, USA) for 10 min, and blocked with phosphate-buffered saline containing 3% bovine serum albumin (GenDEPOT, TX, USA). Thereafter, the oocytes were incubated with a rabbit anti-α-tubulin antibody (1:200; Cell Signaling Technology, MA, USA) and subsequently mounted on slides using VECTASHIELD antifade mounting medium with 4,6-diamidino-2-phenylindole (Vector Laboratories, Peterborough, UK) to visualize the chromosomes using a fluorescence microscope (BX51; Olympus, Tokyo, Japan). Oocytes with well-organized bipolar spindles and tightly aligned chromosomes at metaphase were scored as normal.
RNA sequencing for mRNA expression
Post SM administration, ovaries were collected from the mice post-ovulation, and total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturers’ protocol. The purity and integrity of the extracted RNA were evaluated using a NanoDrop ND-2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and Agilent 2100 bioanalyzer (Agilent Technologies, Amstelveen, The Netherlands). All samples showed high purity (optical density (OD)260/OD280> 1.80) and integrity (RNA integrity number >7.0). Sequencing was performed using the Illumina NextSeq 500 platform. A fold-change value >1.5 and a P-value <0.05 were considered thresholds to identify DEGs.
Validation of selected DEGs in the ovaries
To confirm the mRNA microarray results, validation was performed on significant genes of interest (Fgfr2, Gng8, Rasal1, Rasgrf1, Rasgrf2, NF1, Fgf13, Shc3, and Egf) using real-time qPCR. Complementary DNA (cDNA) was synthesized from the extracted total RNA using iScript cDNA Synthesis kit (Bio-Rad Laboratories, Hercules, CA, USA), according to the manufacturers’ protocol. qPCR was performed in a final reaction volume of 20 μL using a QuantStudio 6 Flex Real-time PCR system with fluorescent probe-based TaqMan assays, according to the manufacturers’ protocol (Thermo Fisher Scientific). The cycle threshold was normalized and compared using glyceraldehyde 3-phosphate dehydrogenase as the internal standard.
Assessment of in vitro developmental competence
Post SM administration, the mice were superovulated with an intraperitoneal injection of 5 IU of PMSG, followed by 5 IU of hCG 48 h later. Superovulated mice were mated with fertile male mice, and the day of vaginal plugging was designated as 0.5 dpc. At 1.5 dpc, 2-cell embryos were collected and cultured up to the blastocyst stage for three days. DNA fragmentation was determined by transferase dUTP nick end labeling (Promega, WI, USA). Blastocyst quality was assessed using the following three parameters: number of blastomeres, DNA fragmentation, and apoptotic index per blastocyst. The apoptotic index was calculated for each blastocyst as follows: apoptotic index= (number of TUNEL-positive nuclei/total number of nuclei) × 100.
Assessment of in vivo implantation potential
Post SM administration, superovulation and mating were achieved as described above. The day of vaginal plugging was designated as 0.5 dpc. At 9.5 dpc, pregnant mice were sacrificed to assess the implantation potential following three parameters: pregnancy rate, total number of litter, and litter weight.
Statistical analysis
Data are presented as mean ± standard error of the mean. The significance of difference between two groups was determined by a Student’s t-test using GraphPad Prism version 8.4.0 (GraphPad Software, La Jolla, CA, USA). The significance of difference between the expected frequencies was determined using Fisher’s exact test. A P-value < 0.05 was considered statistically significant.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Author Contributions
Kim, J: conception, experiments, writing, and revision of the manuscript. You, S: experiments, revision of the manuscript, and correspondence.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This work was supported by a project grant from the Korea Institute of Oriental Medicine (KSN2021240).
References
- 1. Aramillo Irizar P, Schäuble S, Esser D, Groth M, Frahm C, Priebe S, Baumgart M, Hartmann N, Marthandan S, Menzel U, Müller J, Schmidt S, Ast V, et al. Transcriptomic alterations during ageing reflect the shift from cancer to degenerative diseases in the elderly. Nat Commun. 2018; 9:327. https://doi.org/10.1038/s41467-017-02395-2 [PubMed]
- 2. Amanvermez R, Tosun M. An Update on Ovarian Aging and Ovarian Reserve Tests. Int J Fertil Steril. 2016; 9:411–15. https://doi.org/10.22074/ijfs.2015.4591 [PubMed]
- 3. Broekmans FJ, Soules MR, Fauser BC. Ovarian aging: mechanisms and clinical consequences. Endocr Rev. 2009; 30:465–93. https://doi.org/10.1210/er.2009-0006 [PubMed]
- 4. te Velde ER, Pearson PL. The variability of female reproductive ageing. Hum Reprod Update. 2002; 8:141–54. https://doi.org/10.1093/humupd/8.2.141 [PubMed]
- 5. Cimadomo D, Fabozzi G, Vaiarelli A, Ubaldi N, Ubaldi FM, Rienzi L. Impact of Maternal Age on Oocyte and Embryo Competence. Front Endocrinol (Lausanne). 2018; 9:327. https://doi.org/10.3389/fendo.2018.00327 [PubMed]
- 6. Palomba S, Daolio J, Romeo S, Battaglia FA, Marci R, La Sala GB. Lifestyle and fertility: the influence of stress and quality of life on female fertility. Reprod Biol Endocrinol. 2018; 16:113. https://doi.org/10.1186/s12958-018-0434-y [PubMed]
- 7. Cleary-Goldman J, Malone FD, Vidaver J, Ball RH, Nyberg DA, Comstock CH, Saade GR, Eddleman KA, Klugman S, Dugoff L, Timor-Tritsch IE, Craigo SD, Carr SR, et al, and FASTER Consortium. Impact of maternal age on obstetric outcome. Obstet Gynecol. 2005; 105:983–90. https://doi.org/10.1097/01.AOG.0000158118.75532.51 [PubMed]
- 8. Laopaiboon M, Lumbiganon P, Intarut N, Mori R, Ganchimeg T, Vogel JP, Souza JP, Gülmezoglu AM, and WHO Multicountry Survey on Maternal Newborn Health Research Network. Advanced maternal age and pregnancy outcomes: a multicountry assessment. BJOG. 2014 (Suppl 1); 121:49–56. https://doi.org/10.1111/1471-0528.12659 [PubMed]
- 9. Scott RT
Jr , Hofmann GE. Prognostic assessment of ovarian reserve. Fertil Steril. 1995; 63:1–11. [PubMed] - 10. Anderson RA, Wallace WH, Telfer EE. Ovarian tissue cryopreservation for fertility preservation: clinical and research perspectives. Hum Reprod Open. 2017; 2017:hox001. https://doi.org/10.1093/hropen/hox001 [PubMed]
- 11. Ubaldi F, Vaiarelli A, D’Anna R, Rienzi L. Management of poor responders in IVF: is there anything new? Biomed Res Int. 2014; 2014:352098. https://doi.org/10.1155/2014/352098 [PubMed]
- 12. See CJ, McCulloch M, Smikle C, Gao J. Chinese herbal medicine and clomiphene citrate for anovulation: a meta-analysis of randomized controlled trials. J Altern Complement Med. 2011; 17:397–405. https://doi.org/10.1089/acm.2010.0254 [PubMed]
- 13. Zhou K, Zhang J, Xu L, Wu T, Lim CE. Chinese herbal medicine for subfertile women with polycystic ovarian syndrome. Cochrane Database Syst Rev. 2016; 10:CD007535. https://doi.org/10.1002/14651858.CD007535.pub3 [PubMed]
- 14. Kim J, You S. Samul-tang ameliorates oocyte damage due to cyclophosphamide-induced chronic ovarian dysfunction in mice. Sci Rep. 2020; 10:21925. https://doi.org/10.1038/s41598-020-79013-7 [PubMed]
- 15. Nam MN, Lee AY, Sin SM, Goo YM, J CE. Protective effect of Samultang and its four herbal plants against reactive oxygen species in vitro and cellular system. Korean J Agricult Sci. 2019; 46:427–37.
- 16. Lee CJ, Kapelemera AM, Tsai YZ, Lee CT, Xu MY, Wang CC. Evaluating the Therapeutic Efficacy of Si-Wu-Tang Decoction and Concentrated Extract in Follicular Maldevelopment-Related Menstrual Disorders Through Pharmacokinetic/Pharmacodynamic Studies. Front Pharmacol. 2020; 11:1245. https://doi.org/10.3389/fphar.2020.01245 [PubMed]
- 17. Park MJ, Han SE, Kim HJ, Heo JD, Choi HJ, Ha KT, Yang SW, Lee KS, Kim SC, Kim CW, Joo BS. Paeonia lactiflora improves ovarian function and oocyte quality in aged female mice. Anim Reprod. 2020; 17:e20200013. https://doi.org/10.1590/1984-3143-AR2020-0013 [PubMed]
- 18. Bai L, Shi GY, Yang YJ, Chen W, Zhang LF, Qin C. Rehmannia glutinosa exhibits anti-aging effect through maintaining the quiescence and decreasing the senescence of hematopoietic stem cells. Animal Model Exp Med. 2018; 1:194–202. https://doi.org/10.1002/ame2.12034 [PubMed]
- 19. Yoo MA, Song YK, Jang H, Kim DM, Byun SY. Profiling of skin anti-aging related proteins in human dermal fibroblasts by decursin in Angelica gigas Nakai. Korean J Chem Eng. 2011; 28:880–85. https://doi.org/10.1007/s11814-010-0430-1
- 20. Taymor ML. The regulation of follicle growth: some clinical implications in reproductive endocrinology. Fertil Steril. 1996; 65:235–47. https://doi.org/10.1016/s0015-0282(16)58077-7 [PubMed]
- 21. Morey JS, Ryan JC, Van Dolah FM. Microarray validation: factors influencing correlation between oligonucleotide microarrays and real-time PCR. Biol Proced Online. 2006; 8:175–93. https://doi.org/10.1251/bpo126 [PubMed]
- 22. Younis JS. Ovarian aging and implications for fertility female health. Minerva Endocrinol. 2012; 37:41–57. [PubMed]
- 23. American College of Obstetricians and Gynecologists Committee on Gynecologic Practice and Practice Committee. Female age-related fertility decline. Committee Opinion No. 589. Fertil Steril. 2014; 101:633–34. https://doi.org/10.1016/j.fertnstert.2013.12.032 [PubMed]
- 24. Zhang J, Chen Q, Du D, Wu T, Wen J, Wu M, Zhang Y, Yan W, Zhou S, Li Y, Jin Y, Luo A, Wang S. Can ovarian aging be delayed by pharmacological strategies? Aging (Albany NY). 2019; 11:817–32. https://doi.org/10.18632/aging.101784 [PubMed]
- 25. Hayflick L. How and why we age. Exp Gerontol. 1998; 33:639–53. https://doi.org/10.1016/s0531-5565(98)00023-0 [PubMed]
- 26. Shock NW. Ageing: The biology of senescence. J Gerontol. 1964; 19:521–22. https://doi.org/10.1093/geronj/19.4.521
- 27. Lim J, Luderer U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol Reprod. 2011; 84:775–82. https://doi.org/10.1095/biolreprod.110.088583 [PubMed]
- 28. da Silva PF, Schumacher B. DNA damage responses in ageing. Open Biol. 2019; 9:190168. https://doi.org/10.1098/rsob.190168 [PubMed]
- 29. Yu CE, Oshima J, Fu YH, Wijsman EM, Hisama F, Alisch R, Matthews S, Nakura J, Miki T, Ouais S, Martin GM, Mulligan J, Schellenberg GD. Positional cloning of the Werner’s syndrome gene. Science. 1996; 272:258–62. https://doi.org/10.1126/science.272.5259.258 [PubMed]
- 30. Dai Y, But PP, Chan YP, Matsuda H, Kubo M. Antipruritic and antiinflammatory effects of aqueous extract from Si-Wu-Tang. Biol Pharm Bull. 2002; 25:1175–78. https://doi.org/10.1248/bpb.25.1175 [PubMed]
- 31. Chiu HF, Wu YH, Shen YC, Wang SJ, Venkatakrishnan K, Wang CK. Antioxidant and physiological effects of Si-Wu-Tang on skin and liver: a randomized, double-blind, placebo-controlled clinical trial. Chin Med. 2016; 11:30. https://doi.org/10.1186/s13020-016-0102-0 [PubMed]
- 32. Lim JK, Leprivier G. The impact of oncogenic RAS on redox balance and implications for cancer development. Cell Death Dis. 2019; 10:955. https://doi.org/10.1038/s41419-019-2192-y [PubMed]
- 33. Ferro E, Goitre L, Retta SF, Trabalzini L. The Interplay between ROS and Ras GTPases: Physiological and Pathological Implications. J Signal Transduct. 2012; 2012:365769. https://doi.org/10.1155/2012/365769 [PubMed]
- 34. Fan HY, Liu Z, Mullany LK, Richards JS. Consequences of RAS and MAPK activation in the ovary: the good, the bad and the ugly. Mol Cell Endocrinol. 2012; 356:74–79. https://doi.org/10.1016/j.mce.2011.12.005 [PubMed]
- 35. Vigil D, Cherfils J, Rossman KL, Der CJ. Ras superfamily GEFs and GAPs: validated and tractable targets for cancer therapy? Nat Rev Cancer. 2010; 10:842–57. https://doi.org/10.1038/nrc2960 [PubMed]
- 36. Rongo C. Epidermal growth factor and aging: a signaling molecule reveals a new eye opening function. Aging (Albany NY). 2011; 3:896–905. https://doi.org/10.18632/aging.100384 [PubMed]
- 37. Itamochi H, Oumi N, Oishi T, Taniguchi F, Shoji T, Fujiwara H, Sugiyama T, Suzuki M, Kigawa J, Harada T. Fibroblast growth factor receptor 2 is associated with poor overall survival in clear cell carcinoma of the ovary and may be a novel therapeutic approach. Int J Gynecol Cancer. 2015; 25:570–76. https://doi.org/10.1097/IGC.0000000000000414 [PubMed]
- 38. Tsang M, Dawid IB. Promotion and attenuation of FGF signaling through the Ras-MAPK pathway. Sci STKE. 2004; 2004:pe17. https://doi.org/10.1126/stke.2282004pe17 [PubMed]
- 39. Liu Y, Li S, Tao T, Li X, Zhu Q, Liao Y, Ma J, Sun Y, Liu W. Intrafollicular fibroblast growth factor 13 in polycystic ovary syndrome: relationship with androgen levels and oocyte developmental competence. J Ovarian Res. 2018; 11:87. https://doi.org/10.1186/s13048-018-0455-3 [PubMed]
- 40. Bonnet A, Cabau C, Bouchez O, Sarry J, Marsaud N, Foissac S, Woloszyn F, Mulsant P, Mandon-Pepin B. An overview of gene expression dynamics during early ovarian folliculogenesis: specificity of follicular compartments and bi-directional dialog. BMC Genomics. 2013; 14:904. https://doi.org/10.1186/1471-2164-14-904 [PubMed]
- 41. Yang X, Liaw L, Prudovsky I, Brooks PC, Vary C, Oxburgh L, Friesel R. Fibroblast growth factor signaling in the vasculature. Curr Atheroscler Rep. 2015; 17:509. https://doi.org/10.1007/s11883-015-0509-6 [PubMed]
- 42. Badawy A, Wageah A, El Gharib M, Osman EE. Prediction and diagnosis of poor ovarian response: the dilemma. J Reprod Infertil. 2011; 12:241–48. [PubMed]
- 43. Battaglia C, Genazzani AD, Regnani G, Primavera MR, Petraglia F, Volpe A. Perifollicular Doppler flow and follicular fluid vascular endothelial growth factor concentrations in poor responders. Fertil Steril. 2000; 74:809–12. https://doi.org/10.1016/s0015-0282(00)01517-x [PubMed]
- 44. Umehara T, Urabe N, Obata T, Yamaguchi T, Tanaka A, Shimada M. Cutting the ovarian surface improves the responsiveness to exogenous hormonal treatment in aged mice. Reprod Med Biol. 2020; 19:415–24. https://doi.org/10.1002/rmb2.12345 [PubMed]
- 45. Wynn TA, Ramalingam TR. Mechanisms of fibrosis: therapeutic translation for fibrotic disease. Nat Med. 2012; 18:1028–40. https://doi.org/10.1038/nm.2807 [PubMed]
- 46. Mara JN, Zhou LT, Larmore M, Johnson B, Ayiku R, Amargant F, Pritchard MT, Duncan FE. Ovulation and ovarian wound healing are impaired with advanced reproductive age. Aging (Albany NY). 2020; 12:9686–713. https://doi.org/10.18632/aging.103237 [PubMed]
- 47. Kim SM, Kim JS. A Review of Mechanisms of Implantation. Dev Reprod. 2017; 21:351–59. https://doi.org/10.12717/DR.2017.21.4.351 [PubMed]
- 48. Mcclintock MK. Social control of the ovarian cycle and the function of estrous synchrony. Am Zool. 1981; 21:243–56. https://doi.org/10.1093/icb/21.1.243
- 49. Pedersen T, Peters H. Proposal for a classification of oocytes and follicles in the mouse ovary. J Reprod Fertil. 1968; 17:555–57. https://doi.org/10.1530/jrf.0.0170555 [PubMed]