



Introduction
Myocardial ischemia-reperfusion injury (MI/RI) refers to the recovery of blood supply after a period of myocardial ischemia, the weakening of cardiac function, and the aggravation of cardiac tissue damage [1, 2]. With the development of basic research and clinical treatments, coronary artery bypass grafting, percutaneous coronary angioplasty, and intravenous thrombolysis have become widely used in clinical practice [3, 4]. However, MI/RI limits the success rate of these treatments. A series of damaging changes in cardiac function electrophysiology, myocardial ultrastructure, and energy metabolism, and even severe arrhythmia, caused by ischemia can lead to sudden death. Because of the high incidence and mortality of cardiac I/R injury, much attention has been paid to the mechanism and treatment of this kind of disease. Therefore, it is necessary to explore the pathogenesis and treatment of cardiac I/R injury in order to provide a theoretical basis for clinical therapy.
PA is a triterpenoid compound found in Poria cocos, Ganoderma lucidum, and other traditional Chinese medicine, which has a variety of biological and chemical effects [5]. Research has shown that PA could inhibit breast cancer cells metastasis by inhibiting the phosphorylation of phosphatidylinositol transfer protein [6], or inhibit the proliferation, migration, and invasion of gallbladder cancer cells by inhibiting the expression of AKT [7]. Sun et al. found that pachymic acid can significantly inhibit the proliferation of gastric cancer cell line SGC-7901 and induce G0/G1 cell cycle arrest in a concentration-dependent manner [8]. Wen et al. showed that pachymic acid could significantly reduce the proliferation of osteosarcoma cells in a concentration and time-dependent manner and mediated apoptosis in a dose-dependent manner through PTEN/AKT signals deleted on human chromosome 10 and Caspase 3 and 7 activity [9]. Studies have shown that PA prevented the LPS-induced inflammatory and apoptotic response in cardiomyocytes, which may be mediated by inhibition of the ERK1/2 and p38 pathways [10]. However, the function of pachymic acid in cardiac I/R injury has not been reported. Therefore, we would explore the function and potential mechanism of porcine acid in cardiac I/R injury.
Autophagy is widely found in eukaryotic organisms, and it plays a biodegradable role in cells mainly by degrading long-lived proteins or damaged organelles, which is of great help to maintain the stability of the intracellular environment. Autophagy is involved in a variety of physiological and pathological processes, ad plays a vital role in the clearance of apoptotic cells during growth and development and embryonic development. Studies [11–13] have shown that autophagy plays a biaxially regulated role in the process of tissue ischemia-reperfusion injury and is an essential factor in determining cell survival. In response to various stress environments, autophagy can prevent the triggering of the apoptosis pathway by degrading misfolded proteins and damaged organelles, thus playing an essential role in cell protection. Autophagy can be up-regulated under moderate ischemia-reperfusion treatment, which can provide energy for cells. However, if the stimulation is beyond what the cell can withstand, it will cause autophagy to continue to activate, which in turn will lead to the death of the cells involved in autophagy. In the myocardial reperfusion stage, the mechanism of autophagy in cardiomyocytes stimulated by ROS burst and calcium overload is different from that in the ischemic stage. Beclin1 is a critical protein in the production and processing of autophagosomes. Under normal circumstances, Bcl2 binds to the BH3 structure of Beclin1 to inhibit autophagy. Pant et al. found that ROS can up-regulate the expression of Beclin1, ATG5, and LC3II in hepatocellular carcinoma cells through ROS/Akt/mTOR pathway, resulting in up-regulation of autophagy [14]. Yang et al. found that autophagy was significantly decreased after inhibition of Beclin1 expression in the neuronal H/R model. In addition, the use of antioxidants to interfere with H/R could inhibit the up-regulation of Beclin1. Therefore, the up-regulation of Beclin1 caused by the outbreak of ROS during myocardial reperfusion may be an actual cause of autophagy. Excessive up-regulation of autophagy may aggravate cardiomyocyte injury, and it has been found that inhibition of over-up-regulated autophagy can reduce cardiac I/R [15]. However, Liu et al. found that NR4A2, as a transcription factor in the early response of cardiac I/R, can up-regulate autophagy and inhibit apoptosis through NR4A2/P53/BAX pathway and has a protective effect on cardiomyocytes, which proves that autophagy can also inhibit apoptosis [16]. Therefore, autophagy plays a bi-directional role in cardiac I/R.
Here, our research was performed to investigate whether PA prevents cardiomyocytes injury induced by I/R by inhibiting autophagy and apoptosis.
Results
PA inhibits apoptosis in H/R-treated NMVCs
To explore the function of PA on cardiac function in myocadiac I/R, NMVCs were pretreated with PA (1, 3, 5, 10 μg/ml, Figure 1A) for 30 min. Then the NMVCs were then exposed to hypoxia 1h and reoxygenation for 12h. MTT assay was performed to detect cell viability. The results showed that 3, 5, 10 μg/ml PA could inhibit the decreased cell viability induced by H/R (Figure 1B). TUNEL assay performed that PA10 (μg/ml) prevented the apoptosis level in H/R NMVCs (Figure 1C). Calcein-AM/PI staining revealed that PA reduced the dead cells in NMVCs after H/R (Figure 1D). LDH was increased in NMVCs after H/R, while PA partially recovered the LDH level (Figure 1E). Then we assessed the apoptosis-associated protein level in NMVCs. PA blocked the increased level of Cytochrome-c (Cyt-c), and Bax induced by H/R and reversed the decreased level of Bcl2 induced by H/R (Figure 1F). Flow cytometry also confirmed that PA inhibited apoptosis levels in H/R NMVCs (Figure 1G). Taken together, PA could inhibit apoptosis in H/R induced NMVCs.
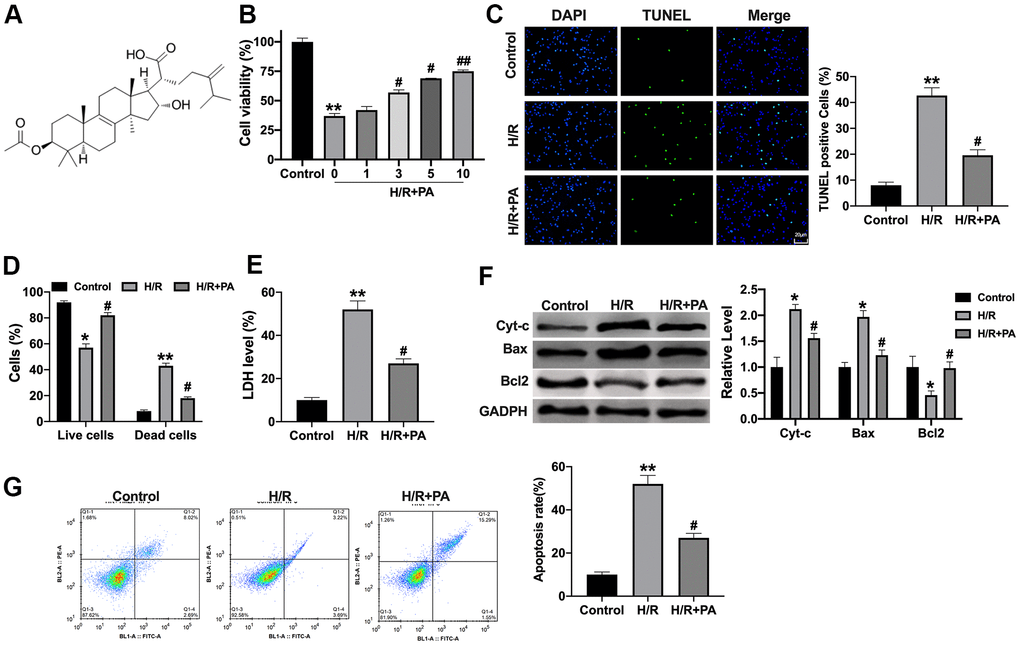
Figure 1. PA inhibits apoptosis in H/R NMVCs. (A) The chemical structure of PA. (B) Cell viability was detected by MTT. n=6, **P<0.01, #P<0.05, ##P<0.01. (C) TUNEL assay was to detect apoptosis level in H/R induced NMVCs treated with 10 μg/ml PA. Scale bar 20 μm. n=4, **P<0.01, #P<0.05. (D) Calcein-AM/PI staining revealed the live cells and dead cells in NMVCs after H/R. n=4, *P<0.05, **P<0.01, #P<0.05. (E) LDH level was determined in NMVCs. n=4, **P<0.01, #P<0.05. (F) The apoptosis-associated protein level in NMVCs, Cytochrome-c (Cyt-c), Bax, and Bcl2. n=4, *P<0.05, #P<0.05. (G) Flow cytometry was used to assess apoptosis level in NMVCs. n=4, **P<0.01, #P<0.05.
PA alleviates mitochondrial function in H/R induced NMVCs
When the NMVCs was hypoxia, the content of ATP was decreased, while PA treated recovered the ATP content (Figure 2A). We also found that PA attenuated ROS production induced by H/R (Figure 2B). The of NMVCs was significantly decreased in the H/R group, while the mPTP opening was recovered in the PA group (Figure 2C). JC-1 staining showed the decreased mitochondrial membrane potential (MMP) in H/R induced NMVCs, while PA-pretreatment reversed the mitochondrial membrane potential (Figure 2D). Meanwhile, PA reversed the GSH and GR activity level in H/R NMVCs (Figure 2E, 2F). In summary, PA could improve the mitochondrial function in H/R induced NMVCs.
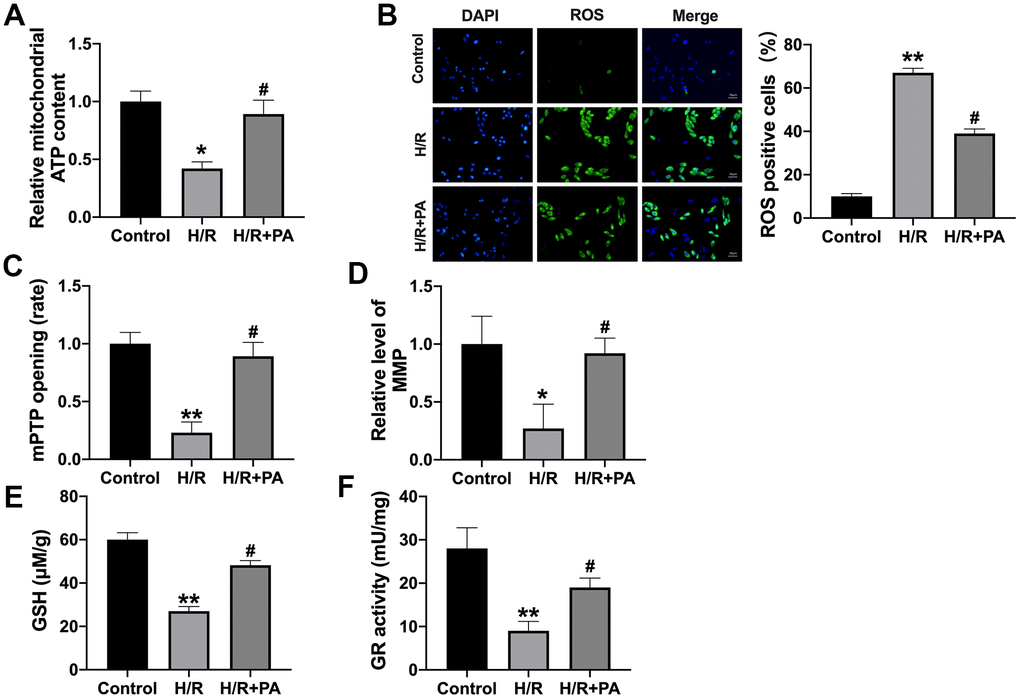
Figure 2. PA improves mitochondrial function in H/R NMVCs. (A) The ATP content was explored in NMVCs. n=4, *P<0.05, #P<0.05. (B) The ROS production in NMVCs was determined. Scale bar 20 μm. n=4, **P<0.01, #P<0.05. (C) The mPTP opening rate in NMVCs was explored. n=5, **P<0.01, #P<0.05. (D) Mitochondrial membrane potential detection in NMVCs. n=4, *P<0.05, #P<0.05. The level of GSH (E) and GR activity (F) in NMVCs. n=4, **P<0.01, #P<0.05.
PA attenuates excessive autophagy in H/R induced NMVCs
Mitochondrial dysfunction is a significant cause of poor prognosis in ischemic heart disease. Mitochondrial autophagy is a crucial process to regulate mitochondrial homeostasis and eliminate damaged mitochondria in time. However, excessive autophagy causes damage to cardiomyocytes. The over-activation of autophagy can promote cell apoptosis, which is the mechanism of injury. We found that excessive autophagy was induced by H/R in NMVCs, which was prevented by PA treatment (Figure 3A–3C). In summary, PA could prevent excessive autophagy in H/R induced NMVCs.
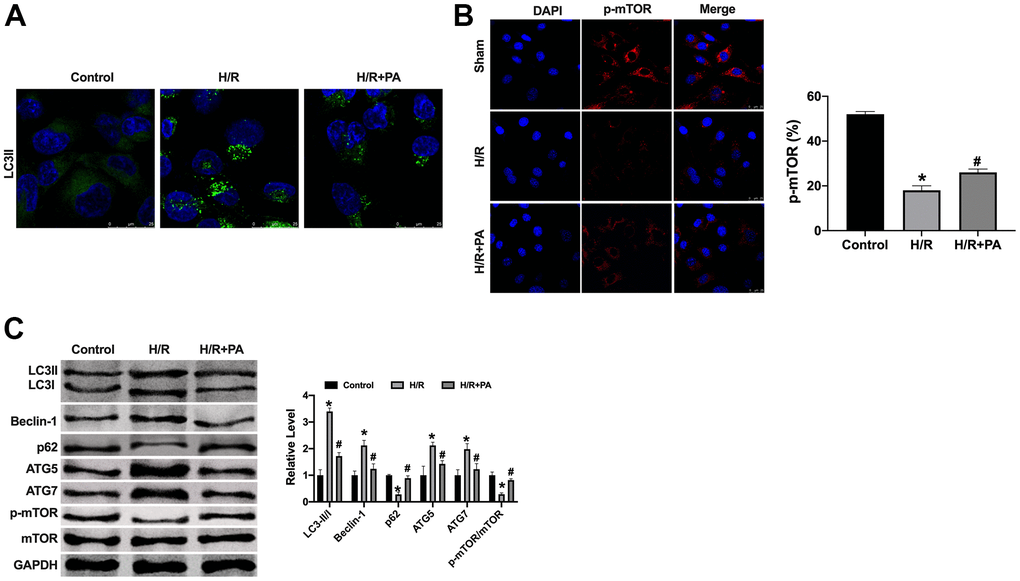
Figure 3. PA inhibited the excessive autophagy in H/R NMVCs. (A, B) Immunofluorescence assay for LC3II and p-mTOR in NMVCs. Scale bar 20 μm. (C) The protein level of LC3, Beclin-1, p62, ATG5, ATG7, mTOR in NMVCs. n=4, *P<0.05, #P<0.05.
PA improves cardiac function in I/R mice
To further explore the PA function on cardiac I/R injury, the I/R model was created in mice. The serum levels of LDH, CK-MB isoenzyme, ROS, MDA, SOD, and GSH-px were assessed by ELISA. PA inhibited the increased level of LDH, CK-MB isoenzyme, ROS, and MDA induced by I/R and recovered the level of SOD and GSH-px in I/R mice (Figure 4A–4F). EF and FS were detected by echocardiography. PA recovered the EF and FS in I/R mice (Figure 5A–5C). Then TTC staining was performed to calculate the infarct size. PA-treated mice revealed a lessened infarct size (Figure 5D). TUNEL assay showed that PA reduced the apoptosis level in I/R mice (Figure 5E). PA reduced the number of inflammatory cells and fibroblasts in the myocardium and reduced the degree of edema of the myocardium and interstitial hyperemia (Figure 5F). The results of the transmission electron microscope (TEM) showed that the mitochondria morphology was swollen and ruptured in the I/R group, while the morphology of mitochondria in the PA group was improved (Figure 5G). Similarly, the apoptosis-associated protein level was consistent with in vitro experiments (Figure 5H). In summary, PA improved apoptosis and autophagy level in I/R mice.
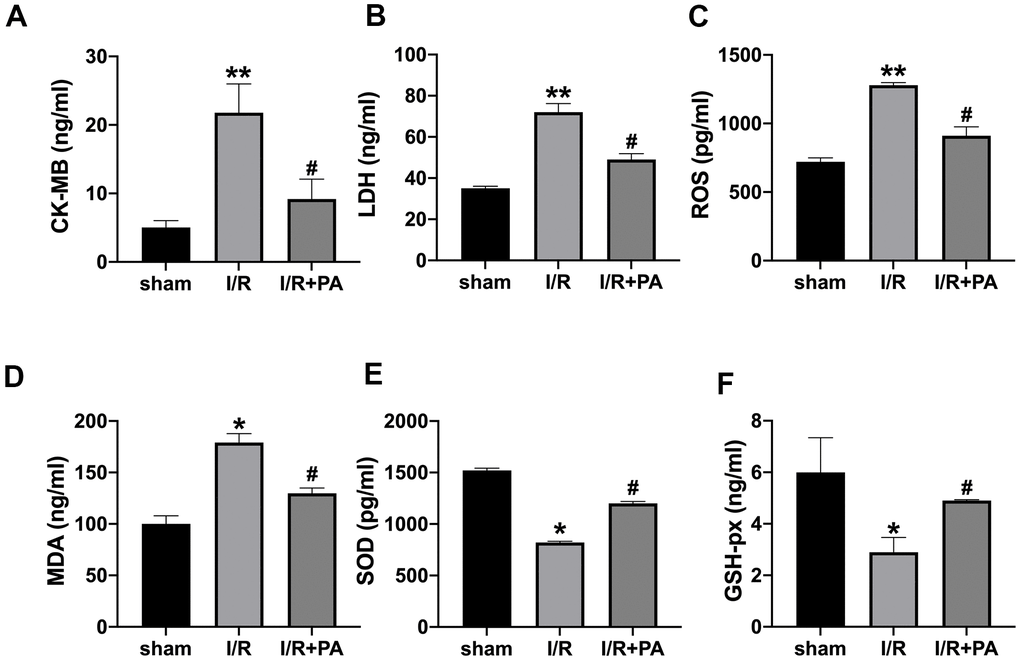
Figure 4. Myocardial injury and oxidative stress indices. Serum CK-MB isoenzyme levels (A), serum LDH levels (B), serum ROS levels (C), serum MDA levels (D), serum SOD levels (E), and serum GSH-px levels (F). n=10, *P<0.05, **P<0.01, #P<0.05.
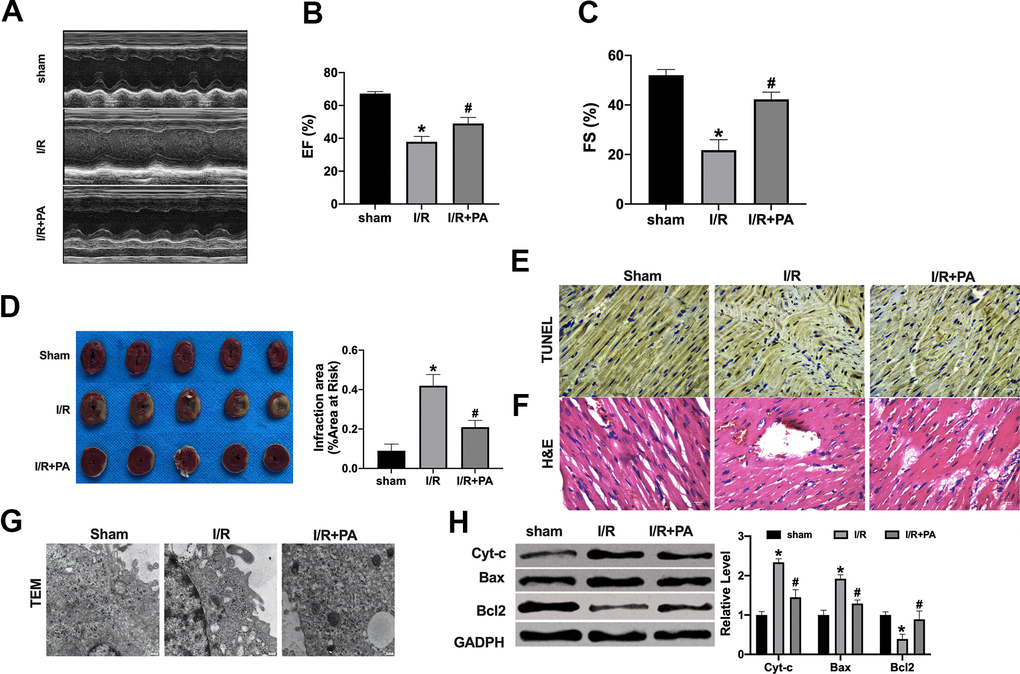
Figure 5. PA ameliorates cardiac function in I/R mice. (A) Cardiac function was detected by echocardiography measurement: EF (B) and FS (C). (D) Infarct sizes as the percent of area at risk (AAR). n=4. *P<0.05, #P<0.05. (E) Representative images of TUNEL staining. (F) Respective HE-stained images of hearts. (G) Transmission electron microscopy was used to detect mitochondrial morphology. (H) The apoptosis-associated protein level in cardiac tissues, Cytochrome-c (Cyt-c), Bax, and Bcl2. n=4, *P<0.05, #P<0.05.
PA inhibited autophagy through the mTOR/p70S6K signal pathway in cardiac I/R injury
The level of autophagy was constantly changing in the stage of I/R. (Figure 6A). Previous research showed that the mTOR/p70S6K signal pathway played a central regulatory role in cell survival, growth and proliferation, and the abnormality of the signal transduction pathway mediated by it is related to a variety of diseases. Here, we assessed the protein level of mTOR and p70S6K in cardiac tissues and cardiomyocytes. Western blot assay performed that PA recovered the protein level of p-mTOR and p-p70S6K in cardiac tissues and cardiomyocytes (Figure 6A, 6B). We detected the autophagy-associated factors in H/R cells after treating them with Rapamycin. We found that Rap co-treated blocked the function of PA on H/R cells (Figure 6C, 6D). Taken together, PA could regulate autophagy via the mTOR/p70S6K signal pathway in cardiac I/R injury.
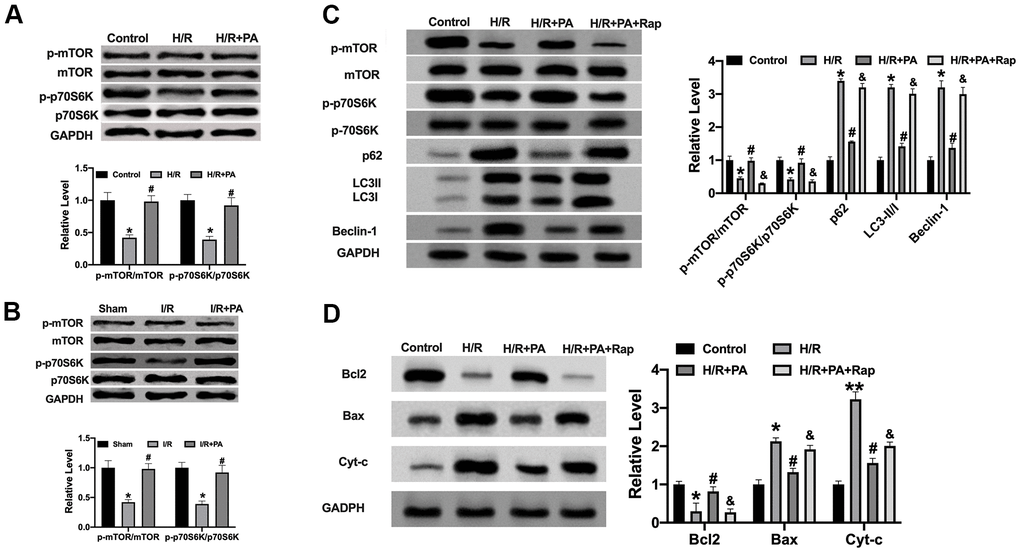
Figure 6. PA regulates autophagy in I/R via mTOR/p70SK6 signal pathway. (A, B) The protein level of mTOR and p70S6K in cardiac tissues and NMVCs. n=4, *P<0.05, #P<0.05. (C) The protein level was detected by Western blot. n=4, *P<0.05 vs. Control group, #P<0.05 vs. H/R group, &P<0.05 vs. H/R+PA group. (D) He apoptosis-associated protein level in cardiac tissues, Cytochrome-c (Cyt-c), Bax, and Bcl2. n=4, P<0.05 vs. Control group, #P<0.05 vs. H/R group, &P<0.05 vs. H/R+PA group.
Discussion
At present, the incidence of acute myocardial infarction in China is increasing year by year. With the development of reperfusion therapy, acute myocardial infarction can be treated effectively in a short period of time. Although it dramatically improves the survival rate of patients with acute myocardial infarction, it also gives rise to new problems [17, 18]. Cardiac I/R injury refers to the oxidative stress injury caused by the second blow caused by HbO2 in the blood when the myocardial infarction criminal blood vessels are opened, and the rapid blood flow flows back into the myocardium when passing through the damaged cardiomyocytes [19]. This injury often leads to further necrosis and apoptosis of cardiomyocytes, mitochondrial dysfunction of viable cardiomyocytes, activation of protein kinases, a large number of inflammatory reactions, which lead to poor long-term prognosis and even death of patients [20]. Therefore, how to improve cardiac I/R injury is a significant problem to be solved in the field of cardiovascular disease.
In recent years, many scholars have studied the effect and mechanism of traditional Chinese medicine on ischemia-reperfusion injury [21]. Salvianolic acid B (Sal B) is the most abundant bioactive component of Salvia miltiorrhiza. Lin et al. found that Sal B improved the infarct size and decreased the levels of serum CK, LDH, and MDA, while SOD significantly increased the level of Sal B, inhibited the expression of Bax, cleaved caspase-9 and cleaved PARP, and promoted the expression of Bcl-2, LC3- II, Beclin1 and VEGF. This shows that Sal B has a cardiovascular protective effect on AMI [22]. Jian J et al. performed that Bauhinia champion flavone inhibits apoptosis and autophagy via the PI3K/Akt pathway in myocardial ischemia-reperfusion injury in rats [23]. Vincent A et al. revealed that MLC901performed acute and long-term cardioprotective effects in cardiac I/R injury [24]. YiXin-Shu reduced cardiac I/R damage by inhibiting mitochondrial-related apoptosis and upregulating liver-X-receptor α [25]. Yan LL et al. performed that Gualou Xiebai Decoction improved cardiac structure and function in I/R mice through controlling RhoA/ROCK signaling pathway [26]. Here, for the first time, we performed the function of PA on cardiac I/R. PA could protect cardiac function via improving mitochondrial function attenuate apoptosis and autophagy level.
During cardiac I/R, the production of free radicals increases due to the disturbance of mitochondrial oxidative phosphorylation, xanthine oxidase catalysis, and neutrophil respiratory explosion, which leads to a series of adverse consequences such as broken membrane lipid components and proteins, which inactivate sarcoplasmic reticulum calcium-dependent ATP enzyme. Meanwhile, ROS in the mitochondria, a large number of species will destroy the mitochondrial membrane structure, resulting in a significant release of cytochrome C, thus initiating apoptosis in the mitochondrial pathway. It has also been found that mitochondria can regulate the original ROS by increasing their own ROS, which is called ROS-induced ROS release (RIRR) [27]. ROS produced by the RIRR pathway can lead to the opening of mPTP, the overload of Ca2+, and the collapse of mitochondrial membrane potential, which further aggravates I/R injury [28]. Therefore, improving the mitochondria function could alleviate the I/R injury. Here, we found that PA recovered the ATP content, MMP, mPTP opening rate, GSH, and GR activity and reduced the ROS production in H/R NMVCs, which performed the improvement on mitochondrial function.
At present, the research on the role of cell autophagy in cardio-cerebrovascular diseases is more and more in-depth. Astragaloside (AS) is not only the main component of Astragalus polysaccharides, but also the most effective component of traditional Chinese medicine Astragalus membranaceus. Yang JJ et al. Astragaloside-IV observed that rescued the H/R injury-induced apoptosis and autophagy in H9c2 cells. AS-IV treatment can stimulate the overexpression of GATA-4 and further enhanced the myocardial protection effect of GATA-4 [29]. Shi X et al. detected that XuefuZhuyu decoction (XFZY) decreased the formation of autophagosomes and downregulated the expression of LC3-II/I ratio and Beclin 1 after H/R. Meanwhile, XFZY could improve cell viability and prevent apoptosis by inhibiting autophagy [30].
Mammalian target of rapamycin (mTOR) is a serine/threonine kinase of the phosphoinositol 3 kinase-related kinase family, which regulates the proliferation, survival, invasion and metastasis of tumor cells by activating ribosomal kinases. The p70S6kinase, p70S6K, is the main effector of mTOR downstream of the mTOR/ p70S6K signaling pathway. The mTOR/ p70S6K signaling pathway regulates autophagy in multiple cellular processes. In our research, we found that PA could inhibit the expression level of LC3II by preventing the mTOR/P70S6K signal pathway.
Conclusions
We revealed that PA prevented cardiac I/R injury by blocking apoptosis and autophagy in vivo and vitro, mTOR/p70S6K pathway may involve in this progression, which provides a new basis for clinical application.
Materials and Methods
Animal model
30 SPF grade of C57BL/6 mice, weighing (22 ±1) g. I/R model was created as follows [15]. The skin was prepared and disinfected in the operation area of the I/R model, the skin was cut between the third and fourth ribs on the left side of the sternum, the subcutaneous tissue and muscle were bluntly separated layer by layer, the chest cavity was opened, the heart was fully exposed with a chest opener, and the LAD was carefully identified under stereoscopic microscope. The suture needle with 6-0 thread was used to pass through the surface layer of the myocardium at the inferior edge of the auricle 1mm, and the LAD was ligated under pressure. After ligation of LAD, it was found that the anterior wall of the left ventricle was whitened, the local contractile movement was limited, the left atrial appendage was filled and inflated, and the ST segment was continuously arched and raised, indicating the success of myocardial ischemia. The chest cavity was closed by simple suture, and the chest cavity was opened again after ischemia for 30 min. The polyethylene tube was removed, and the ligation line was cut. The surface of the heart changed from pallor to red, and the elevated part of the ST segment of ECG fell back to judge the success of reperfusion. When the mice wake up, extubate the trachea and put it on a 37° C blanket for the night. After 12 hours, the heart function of mice was detected by echocardiography, and the heart was separated and stored at-80° C.
The PA-treated mice were given suspensions by gavage containing PA (5 mg/kg body weight) for 3 days before the model was induced, Then the mice were constructed for the I/R model.
All animal work has been reviewed and approved by the Animal Care and Use Committee our hospital and carried out in accordance with the National Institutes of Health guide for the care and use of laboratory animals.
NMVCs cultured
C57BL/6 mice (1-3 days old) were selected, soaked in alcohol for about 3 min, thoracotomy was performed to remove the apex of the heart under sterile conditions, and pre-cooled with PBS at 4° C for 3-4 times to remove residual blood. Then the heart was cut into a size of about 1 mm3, mixed digestive juices of 0.25% trypsin were added, and digested in a water bath at 37° C. After the first digestion, natural precipitation was required, the supernatant was discarded, and the precipitation was taken for digestion. After natural precipitation, the supernatant was taken for digestion. DMEM containing 10% FBS was added to terminate digestion. After repeated digestion for 5-6 times, all the collected supernatant was filtered with a 200-mesh stainless steel screen to remove the residual myocardial tissue mass, and the cell suspension was collected. After centrifugation for 5 min at 1000 r/min, the supernatant was discarded, and the cells were suspended once in DMEM medium containing 10%FBS. The supernatant was centrifuged at 1000 r/min, and the cells were resuspended and inoculated in a 100 mm culture plate. The cells were adhered to the wall at differential speed for 60min. The supernatant was absorbed and replanted to the plate, which was the primary cultured cardiomyocytes. In order to inhibit the growth of fibroblasts, Brdu with a final concentration of 0.1 mmol/L was added to the medium, and the solution was changed after 24 h culture, and then once every 2 days. Cells from the third day of culture were collected for subsequent experiments.
NMVCs were pretreated with PA (MEC, USA) for 1, 3, 5, 10 μg/ml for 30min. Then the NMVCs were then exposed to hypoxia 1h (0.1% O2, 5% CO2) and reoxygenation for 12h.
Transmission electron microscope (TEM)
The left ventricular tissues were fixed with glutaraldehyde and Osmic acid, dehydrated step by step with ethanol acetone, embedded with epoxy resin, ultra-thin section of LKB- V type ultrathin section mechanism, ultra-thin section, double staining of uranyl acetate and lead citrate, and autophagy bodies in cardiomyocytes were observed and counted under TEM.
Western blot
The cardiac tissues were extracted by RIPA 200 μL, and the protein level of the sample was detected by BCA method. After balancing the protein concentration of each group, the protein was separated by SDS-PAGE gel electrophoresis. The protein was transferred to NC membrane by wet method, 5% skimmed milk powder was incubated for 2h, primary antibody was added and incubated overnight in a closed bag at 4° C, and the fluorescent second antibody was incubated at room temperature for 1 h. The gray level of the target strip is analyzed, and the result is expressed by the ratio of the target to the loading control. The primary antibody was shown as follow. Bax Polyclonal Antibody (Proteintech, 50599-2-Ig, 21kDa); Bcl2 Polyclonal Antibody (Proteintech, 12789-1-AP, 25kDa); Cytochrome C Polyclonal Antibody (Proteintech, 10993-1-AP, 12kDa); LC3 Polyclonal Antibody (Proteintech, 14600-1-AP, 15-20kDa); Beclin 1 Polyclonal Antibody (Proteintech, 11306-1-AP, 52kDa); P62/SQSTM1 Polyclonal Antibody (Proteintech, 18420-1-AP, 62kDa); ATG5 Polyclonal Antibody (Proteintech, 18420-1-AP, 52kDa); ATG7 Polyclonal Antibody (Proteintech, 10088-2-AP, 78kDa); Phospho-mTOR (Ser2448) Monoclonal Antibody (Proteintech, 67778-1-Ig, 289kDa); mTOR Monoclonal Antibody (Proteintech, 66888-1-Ig, 289kDa); P70(S6K) Polyclonal Antibody (Proteintech, 14485-1-AP, 70kDa); Phospho-p70 S6 Kinase (Thr389/Thr412) Antibody (Affinit, AF3228, 70kDa); GAPDH Monoclonal Antibody (Proteintech, 60004-1-Ig, 36kDa).
TUNEL assay
Left ventricular tissues or NMVCs were fixed with 4% paraformaldehyde, ethanol gradient dehydration, paraffin-embedded and sectioned, operated according to TUNEL kit, and cardiomyocyte apoptosis was observed by fluorescence microscope. The specific experimental operation refers to the instruction manual of DeadEed™Fluorometric TUNEL detection kit.
ROS kit assay
The cells were inoculated into 24-well plates, and after treatment, the cells were cleaned 3 times with PBS preheated at 37° C for 5 min each time. DCFH-DA was diluted with PBS buffer (1:1000), and working solution was prepared. 200 μL working solution was added to each well, and incubated in the incubator against light for 30 min. Then the incubator was cleaned with PBS at 37° C for 3 times, 5 min each time, and then observed and photographed under a fluorescence microscope. The image-Pro Plus software was used to analyze the fluorescence intensity, and the average fluorescence intensity represented the relative ROS level.
Apoptosis analysis
The cells were inoculated in 24-well plates with a density of 5×104 cells per well. After treatment, the cells were fixed with poformaldehyde and permeated with 0.01%Triton X-100 (Sigma, USA). The cells were incubated by TUNEL enzyme reaction mixture for 60 min, and then the cells were washed with PBS for 3 times and observed under a fluorescence microscope.
Echocardiography
Mice were anesthetized by isoflurane, the left interventricular septum thickness (IVS) and the left interventricular septum thickness (IVS) were measured by high resolution small animal ultrasound imaging system. S), left ventricular posterior wall end-diastolic thickness (LVPW, d), LVPW (S), ejection fraction, EF), each index was measured and averaged over 2-3 cardiac cycles.
Statistical analysis
The data were analyzed by GraphPad 8.0. the experimental data were expressed in ±SEM, and the difference was statistically significant by T-test or one-way ANOVA. The difference was statistically significant (P < 0.05).
Author Contributions
Chang Sun and Zhijun Sun contributed to the conception and design of the work, conducted experiments. Na Xi and Xinxin Zhang contributed to the acquisition, analysis, and interpretation of data for the work. Xiaowei Wang drafted the manuscript. Huiyang Cao and Xiaowei Jia critically revised the manuscript. All gave final approval and agreed to be accountable for all aspects of work ensuring integrity and accuracy.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- 1. Ibáñez B, Heusch G, Ovize M, Van de Werf F. Evolving therapies for myocardial ischemia/reperfusion injury. J Am Coll Cardiol. 2015; 65:1454–71. https://doi.org/10.1016/j.jacc.2015.02.032 [PubMed]
- 2. Murphy E, Steenbergen C. Mechanisms underlying acute protection from cardiac ischemia-reperfusion injury. Physiol Rev. 2008; 88:581–609. https://doi.org/10.1152/physrev.00024.2007 [PubMed]
- 3. Davidson SM, Ferdinandy P, Andreadou I, Bøtker HE, Heusch G, Ibáñez B, Ovize M, Schulz R, Yellon DM, Hausenloy DJ, Garcia-Dorado D, and CARDIOPROTECTION COST Action (CA16225). Multitarget Strategies to Reduce Myocardial Ischemia/Reperfusion Injury: JACC Review Topic of the Week. J Am Coll Cardiol. 2019; 73:89–99. https://doi.org/10.1016/j.jacc.2018.09.086 [PubMed]
- 4. Del Re DP, Amgalan D, Linkermann A, Liu Q, Kitsis RN. Fundamental Mechanisms of Regulated Cell Death and Implications for Heart Disease. Physiol Rev. 2019; 99:1765–817. https://doi.org/10.1152/physrev.00022.2018 [PubMed]
- 5. Lee SG, Kim MM. Pachymic acid promotes induction of autophagy related to IGF-1 signaling pathway in WI-38 cells. Phytomedicine. 2017; 36:82–7. https://doi.org/10.1016/j.phymed.2017.09.020 [PubMed]
- 6. Hong R, Shen MH, Xie XH, Ruan SM. Inhibition of breast cancer metastasis via PITPNM3 by pachymic acid. Asian Pac J Cancer Prev. 2012; 13:1877–80. https://doi.org/10.7314/apjcp.2012.13.5.1877 [PubMed]
- 7. Chen Y, Lian P, Liu Y, Xu K. Pachymic acid inhibits tumorigenesis in gallbladder carcinoma cells. Int J Clin Exp Med. 2015; 8:17781–8. [PubMed]
- 8. Sun KX, Xia HW. Pachymic acid inhibits growth and induces cell cycle arrest and apoptosis in gastric cancer SGC-7901 cells. Oncol Lett. 2018; 16:2517–24. https://doi.org/10.3892/ol.2018.8899 [PubMed]
- 9. Wen H, Wu Z, Hu H, Wu Y, Yang G, Lu J, Yang G, Guo G, Dong Q. The anti-tumor effect of pachymic acid on osteosarcoma cells by inducing PTEN and Caspase 3/7-dependent apoptosis. J Nat Med. 2018; 72:57–63. https://doi.org/10.1007/s11418-017-1117-2 [PubMed]
- 10. Li FF, Yuan Y, Liu Y, Wu QQ, Jiao R, Yang Z, Zhou MQ, Tang QZ. Pachymic acid protects H9c2 cardiomyocytes from lipopolysaccharide-induced inflammation and apoptosis by inhibiting the extracellular signal-regulated kinase 1/2 and p38 pathways. Mol Med Rep. 2015; 12:2807–13. https://doi.org/10.3892/mmr.2015.3712 [PubMed]
- 11. Heusch G. Myocardial ischaemia-reperfusion injury and cardioprotection in perspective. Nat Rev Cardiol. 2020; 17:773–89. https://doi.org/10.1038/s41569-020-0403-y [PubMed]
- 12. Dong Y, Chen H, Gao J, Liu Y, Li J, Wang J. Molecular machinery and interplay of apoptosis and autophagy in coronary heart disease. J Mol Cell Cardiol. 2019; 136:27–41. https://doi.org/10.1016/j.yjmcc.2019.09.001 [PubMed]
- 13. Lu LQ, Tian J, Luo XJ, Peng J. Targeting the pathways of regulated necrosis: a potential strategy for alleviation of cardio-cerebrovascular injury. Cell Mol Life Sci. 2021; 78:63–78. https://doi.org/10.1007/s00018-020-03587-8 [PubMed]
- 14. Pant K, Saraya A, Venugopal SK. Oxidative stress plays a key role in butyrate-mediated autophagy via Akt/mTOR pathway in hepatoma cells. Chem Biol Interact. 2017; 273:99–106. https://doi.org/10.1016/j.cbi.2017.06.001 [PubMed]
- 15. Fan Q, Tao R, Zhang H, Xie H, Lu L, Wang T, Su M, Hu J, Zhang Q, Chen Q, Iwakura Y, Shen W, Zhang R, Yan X. Dectin-1 Contributes to Myocardial Ischemia/Reperfusion Injury by Regulating Macrophage Polarization and Neutrophil Infiltration. Circulation. 2019; 139:663–78. https://doi.org/10.1161/CIRCULATIONAHA.118.036044 [PubMed]
- 16. Liu H, Liu P, Shi X, Yin D, Zhao J. NR4A2 protects cardiomyocytes against myocardial infarction injury by promoting autophagy. Cell Death Discov. 2018; 4:27. https://doi.org/10.1038/s41420-017-0011-8 [PubMed]
- 17. Yellon DM, Hausenloy DJ. Myocardial reperfusion injury. N Engl J Med. 2007; 357:1121–35. https://doi.org/10.1056/NEJMra071667 [PubMed]
- 18. Hausenloy DJ, Yellon DM. Targeting Myocardial Reperfusion Injury--The Search Continues. N Engl J Med. 2015; 373:1073–5. https://doi.org/10.1056/NEJMe1509718 [PubMed]
- 19. Hausenloy DJ, Yellon DM. Myocardial ischemia-reperfusion injury: a neglected therapeutic target. J Clin Invest. 2013; 123:92–100. https://doi.org/10.1172/JCI62874 [PubMed]
- 20. Lejay A, Fang F, John R, Van JA, Barr M, Thaveau F, Chakfe N, Geny B, Scholey JW. Ischemia reperfusion injury, ischemic conditioning and diabetes mellitus. J Mol Cell Cardiol. 2016; 91:11–22. https://doi.org/10.1016/j.yjmcc.2015.12.020 [PubMed]
- 21. Han JY, Li Q, Ma ZZ, Fan JY. Effects and mechanisms of compound Chinese medicine and major ingredients on microcirculatory dysfunction and organ injury induced by ischemia/reperfusion. Pharmacol Ther. 2017; 177:146–73. https://doi.org/10.1016/j.pharmthera.2017.03.005 [PubMed]
- 22. Lin C, Liu Z, Lu Y, Yao Y, Zhang Y, Ma Z, Kuai M, Sun X, Sun S, Jing Y, Yu L, Li Y, Zhang Q, Bian H. Cardioprotective effect of Salvianolic acid B on acute myocardial infarction by promoting autophagy and neovascularization and inhibiting apoptosis. J Pharm Pharmacol. 2016; 68:941–52. https://doi.org/10.1111/jphp.12567 [PubMed]
- 23. Jian J, Xuan F, Qin F, Huang R. Bauhinia championii flavone inhibits apoptosis and autophagy via the PI3K/Akt pathway in myocardial ischemia/reperfusion injury in rats. Drug Des Devel Ther. 2015; 9:5933–45. https://doi.org/10.2147/DDDT.S92549 [PubMed]
- 24. Vincent A, Covinhes A, Barrère C, Gallot L, Thoumala S, Piot C, Heurteaux C, Lazdunski M, Nargeot J, Barrère-Lemaire S. Acute and long-term cardioprotective effects of the Traditional Chinese Medicine MLC901 against myocardial ischemia-reperfusion injury in mice. Sci Rep. 2017; 7:14701. https://doi.org/10.1038/s41598-017-14822-x [PubMed]
- 25. Zhao Y, Xu L, Qiao Z, Gao L, Ding S, Ying X, Su Y, Lin N, He B, Pu J. YiXin-Shu, a ShengMai-San-based traditional Chinese medicine formula, attenuates myocardial ischemia/reperfusion injury by suppressing mitochondrial mediated apoptosis and upregulating liver-X-receptor α. Sci Rep. 2016; 6:23025. https://doi.org/10.1038/srep23025 [PubMed]
- 26. Yan LL, Zhang WY, Wei XH, Yan L, Pan CS, Yu Y, Fan JY, Liu YY, Zhou H, Han JY, Yao XS. Gualou Xiebai Decoction, a Traditional Chinese Medicine, Prevents Cardiac Reperfusion Injury of Hyperlipidemia Rat via Energy Modulation. Front Physiol. 2018; 9:296. https://doi.org/10.3389/fphys.2018.00296 [PubMed]
- 27. Zorov DB, Filburn CR, Klotz LO, Zweier JL, Sollott SJ. Reactive oxygen species (ROS)-induced ROS release: a new phenomenon accompanying induction of the mitochondrial permeability transition in cardiac myocytes. J Exp Med. 2000; 192:1001–14. https://doi.org/10.1084/jem.192.7.1001 [PubMed]
- 28. Wang J, Toan S, Zhou H. New insights into the role of mitochondria in cardiac microvascular ischemia/reperfusion injury. Angiogenesis. 2020; 23:299–314. https://doi.org/10.1007/s10456-020-09720-2 [PubMed]
- 29. Yang JJ, Zhang XH, Ma XH, Duan WJ, Xu NG, Chen YJ, Liang L. Astragaloside IV enhances GATA-4 mediated myocardial protection effect in hypoxia/reoxygenation injured H9c2 cells. Nutr Metab Cardiovasc Dis. 2020; 30:829–42. https://doi.org/10.1016/j.numecd.2020.01.009 [PubMed]
- 30. Shi X, Zhu H, Zhang Y, Zhou M, Tang D, Zhang H. XuefuZhuyu decoction protected cardiomyocytes against hypoxia/reoxygenation injury by inhibiting autophagy. BMC Complement Altern Med. 2017; 17:325. https://doi.org/10.1186/s12906-017-1822-0 [PubMed]