




Introduction
Non-small cell lung cancer (NSCLC) serves as one of the most prevalent malignancies. It is a primary reason for tumor-induced mortality globally, according to the latest annual statistics report of global cancer [1, 2]. At present, surgical resection is the only acceptable strategy for NSCLC treatment. However, many NSCLC patients are diagnosed at advanced stages, which is not suitable for surgical operation [3]. Unfortunately, the 5-year survival rate remains poor in NSCLC cases, and the recurrence frequency in the patients is high because of drug-resistance or tumor metastasis [4, 5]. Accordingly, timely diagnosis and appropriate treatment are crucial for improving treatment outcomes of NSCLC patients [6]. Therefore, the development of more practical drug candidates and therapeutic strategy is urgently needed.
Increasing evidence has emerged that local anesthetics are able to restrain cancer progression [7]. For example, at the tissue level, the treatment of a range of concentration of local anesthetics, local or topical, has been determined to show a significant inhibitory impact on the epidermal growth factor receptor (EGFR) activation, which serves as a crucial target in cancer cells [8]. Levobupivacaine is a broadly applied long-acting local anesthetic registered for epidural, intrathecal, and ophthalmic anesthesia, infiltration, and nerve block [9]. It has been identified that levobupivacaine pharmacologically presents an anti-tumor function in residual cancer cells [10]. Moreover, several investigations demonstrate that levobupivacaine attenuates cancer progression at a range of specific concentrations in many cancer models, such as prostate and breast cancer [10, 11]. Nevertheless, the influence of levobupivacaine on NSCLC tumorigenesis is still elusive.
Regulated cell death (RCD) shows a significant function in conventional homeostasis preservation and progress in multicellular animals [12]. Ferroptosis, as a distinctive kind of RCD, is initially recognized while a small molecule termed erastin can constrain the potential of triggering an RCD process, particularly in RAS-mutated cancer cells [13]. Ferroptosis is initially issued as an iron-related and non-apoptotic type of oxidative cell disruption in 2012 [14]. RAS-selective lethal small molecular-3 (RSL-3) has presented comparable qualities as erastin [15], discovering this process is featured by the iron-related unusual increase of reactive oxygen species. It is genetically, morphologically, and biochemically distinguished from autophagy, necrosis, and apoptosis [16]. The introduction of ferroptosis causes the interference of cancer progression, and the ferroptosis stimulator repressing tumor growth can be served as potential anti-tumor treatments [17]. Meanwhile, ferroptosis also plays essential roles during pathogenesis of NSCLC [18]. Moreover, tumor suppressor p53 is a ferroptosis inducer in cancer pathogenesis [19]. However, the correlation of levobupivacaine with ferroptosis and p53 in the development of NSCLC is still elusive.
In this study, we focused on the investigation of the impact and the underlying mechanism of levobupivacaine on the development of NSCLC. We identified a novel anti-tumor function of levobupivacaine in NSCLC progression by activating ferroptosis through up-regulating and activating p53.
Results
Levobupivacaine inhibits proliferation and promotes apoptosis of NSCLC cells
To assess the potential function of levobupivacaine in the regulation of NSCLC progression, the A549 and A427 cells were treated with levobupivacaine. MTT assays revealed that the treatment of levobupivacaine decreased the cell viability of the A549 and A427 cells (Figure 1A and 1B). Similarly, the colony formation was decreased by the levobupivacaine treatment in the A549 and A427 cells (Figure 1C and 1D). Furthermore, cell apoptosis was increased in the levobupivacaine-treated cells (Figure 1E and 1F). Moreover, the G0/G1 phase cells were enhanced while the S phase cells were reduced by levobupivacaine (Figure 1G and 1H), suggesting that levobupivacaine was able to promote G0/G1 cell cycle arrest in the cells. Together our data indicate that levobupivacaine can inhibit proliferation and promote apoptosis of NSCLC cells.
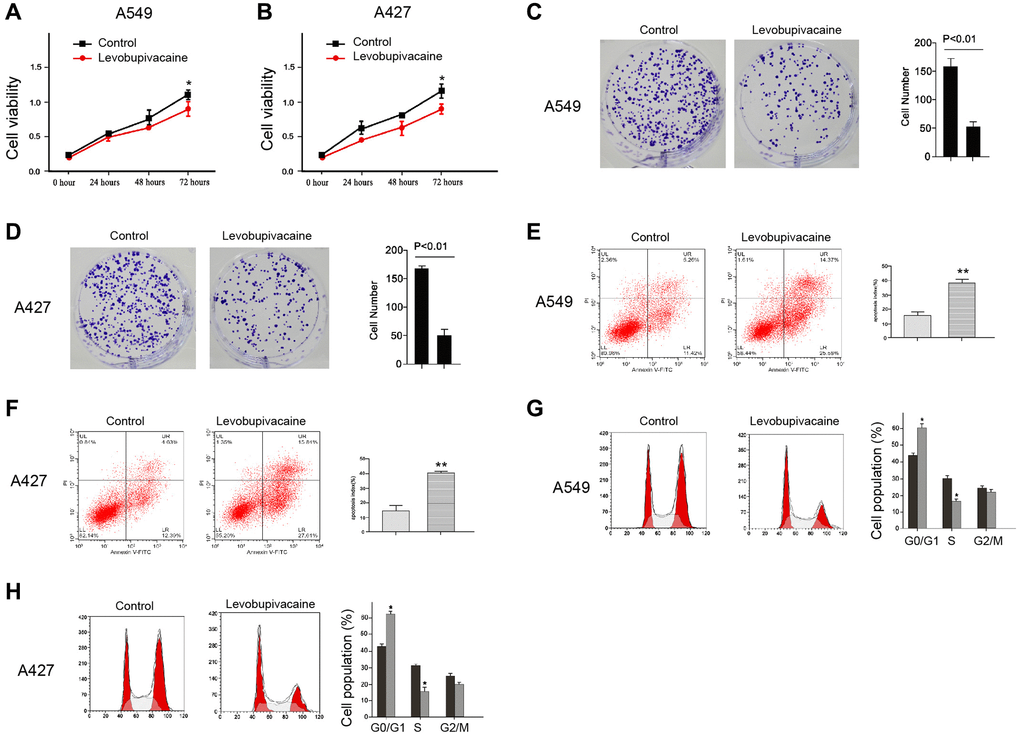
Figure 1. Levobupivacaine inhibits proliferation and promotes apoptosis of NSCLC cells. (A–E) The A549 and A427 cells were treated with levobupivacaine (2 mM) or equal volume saline. (A and B) The cell viability was analyzed by the MTT assays in the cells. (C and D) The cell proliferation was measured by the colony formation assays in the cells. (E and F) The cell apoptosis was measured by flow cytometry analysis in the cells. (G and H) The cell cycle was analyzed by flow cytometry analysis in the cells. Data are presented as mean ± SD. Statistic significant differences were indicated: *P < 0.05, **P < 0.01.
Levobupivacaine represses invasion and migration of NSCLC cells
Next, we determined the function of levobupivacaine in regulating the migration and invasion of NSCLC cells. Transwell assays revealed that the migration and invasion were remarkably inhibited by the treatment of levobupivacaine in A549 and A427 cells (Figure 2A and 2B). Similarly, levobupivacaine significantly reduced the wound healing proportion in the cells (Figure 2C and 2D), suggesting that levobupivacaine is able to inhibit NSCLC cell migration and invasion.
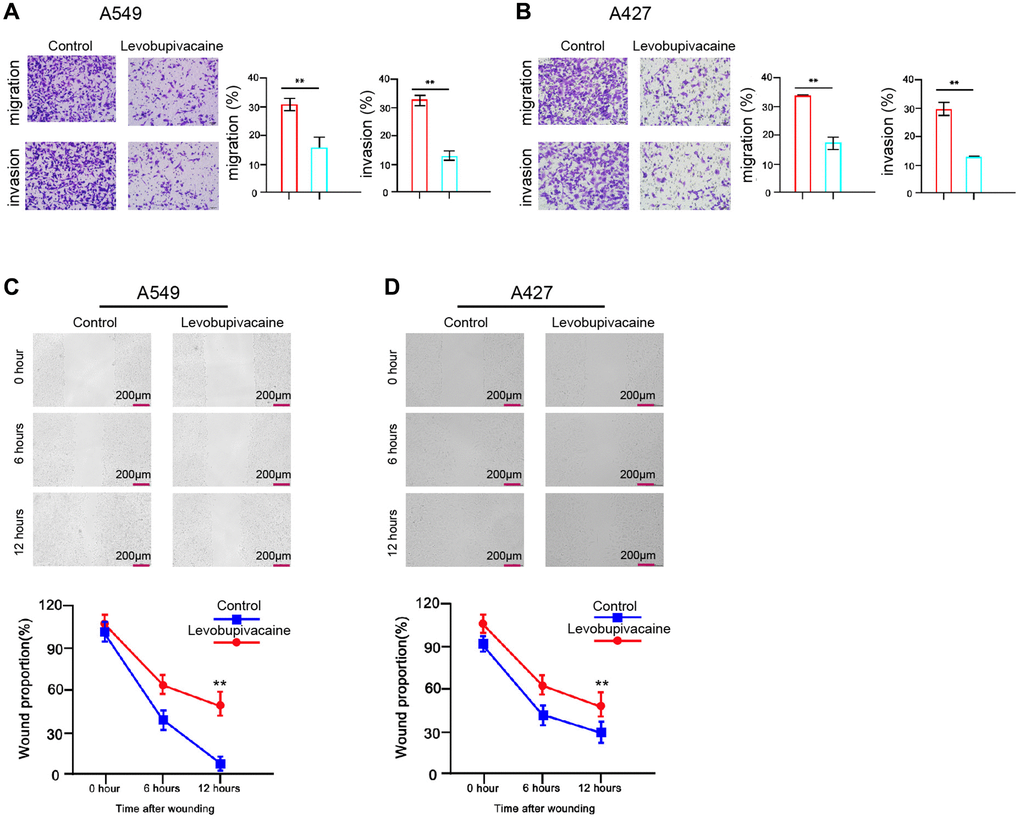
Figure 2. Levobupivacaine represses invasion and migration of NSCLC cells. (A–D) The A549 and A427 cells were treated with levobupivacaine (2 mM) or equal volume saline. (A and B) The cell migration and invasion were examined by transwell assays in the cells. (C and D) The migration and invasion were measured by wound healing assays in the cells. The wound healing proportion was shown. Data are presented as mean ± SD. Statistic significant differences were indicated: **P < 0.01.
Levobupivacaine induces ferroptosis of NSCLC cells
To assess the role of levobupivacaine in ferroptosis, we analyzed the effect of levobupivacaine on the erastin-induced inhibition of cell growth and the intracellular levels of reactive oxygen species, iron, and Fe2+, and the expression of GPX4 and SLC7A11, which was the surrogate markers for ferroptosis. Significantly, the treatment of levobupivacaine enhanced the erastin-induced inhibition of cell growth in the A549 and A427 cells, in which erastin served as the activator of ferroptosis (Figure 3A and 3B). The effect of co-treatment of erastin and Ferrostatin-1 served as the negative control (Figure 3A and 3B). Meanwhile, the treatment of levobupivacaine remarkably increased the levels of ROS in the A549 and A427 cells (Figure 3C). The iron levels were enhanced by levobupivacaine in the cells (Figure 3D). Besides, the treatment of levobupivacaine promoted the accumulation of Fe2+ in the cells (Figure 3E). Moreover, the expression of GPX4 and SLC7A11 was inhibited by levobupivacaine in the cells (Figure 3F and 3G). The effect of erastin on the ferroptosis markers was the positive control in the system (Figure 3C–3G). Taken together, levobupivacaine can induce ferroptosis in NSCLC cells.
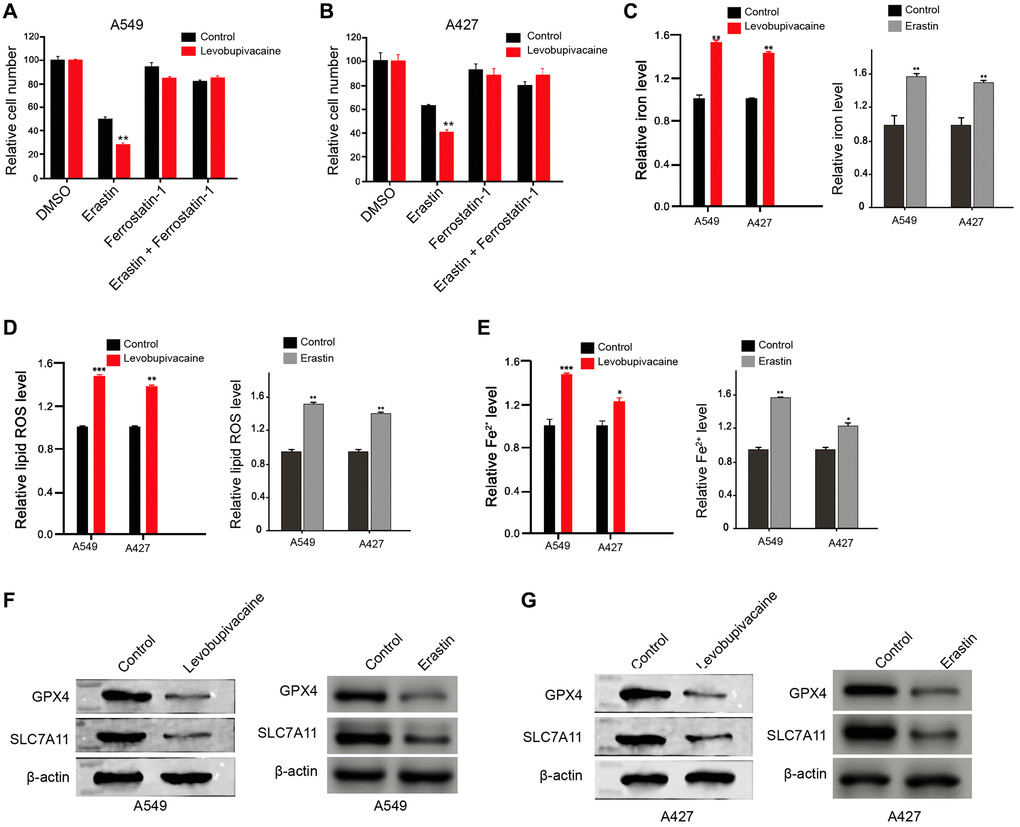
Figure 3. Levobupivacaine induces ferroptosis in NSCLC cells. (A and B) The A549 and A427 cells were co-treated with 5 mmol/L erastin or ferrostatin (1 mmol/L) and 2 mM levobupivacaine. The cell growth was analyzed by MTT assays. (C–G) The A549 and A427 cells were treated with levobupivacaine (2 mM) or equal volume saline. The erastin was used as a positive control. (C) The levels of ROS were measure by flow cytometry analysis in the cells. (D and E) The levels of iron and Fe2+ were analyzed by Iron Assay Kit. (F and G) The expression of GPX4, SLC7A11, and β-actin was measured by Western blot analysis in the cells. Data are presented as mean ± SD. Statistic significant differences were indicated: *P < 0.05, **P < 0.01.
Levobupivacaine up-regulates and activates p53 in NSCLC cells
Then, we further explored the mechanism of levobupivacaine-mediated NSCLC progression. Significantly, we identified that the treatment of levobupivacaine enhanced the total expression of p53 and nucleus accumulation in the A549 and A427 cells (Figure 4A and 4B). The p53 overexpression efficiency was confirmed in the cells (Figure 4C). We validated that the overexpression of p53 increased the erastin-induced inhibition of cell growth in the A549 and A427 cells (Figure 4D and 4E). Meanwhile, p53 overexpression remarkably induced the levels of ROS, iron, and Fe2+ in A549 and A427 cells (Figure 4F–4H). Moreover, the expression of GPX4 and SLC7A11 was inhibited by p53 overexpression in the cells (Figure 4I and 4J). Together these data indicate that levobupivacaine activates p53 and p53 induces ferroptosis in NSCLC cells.
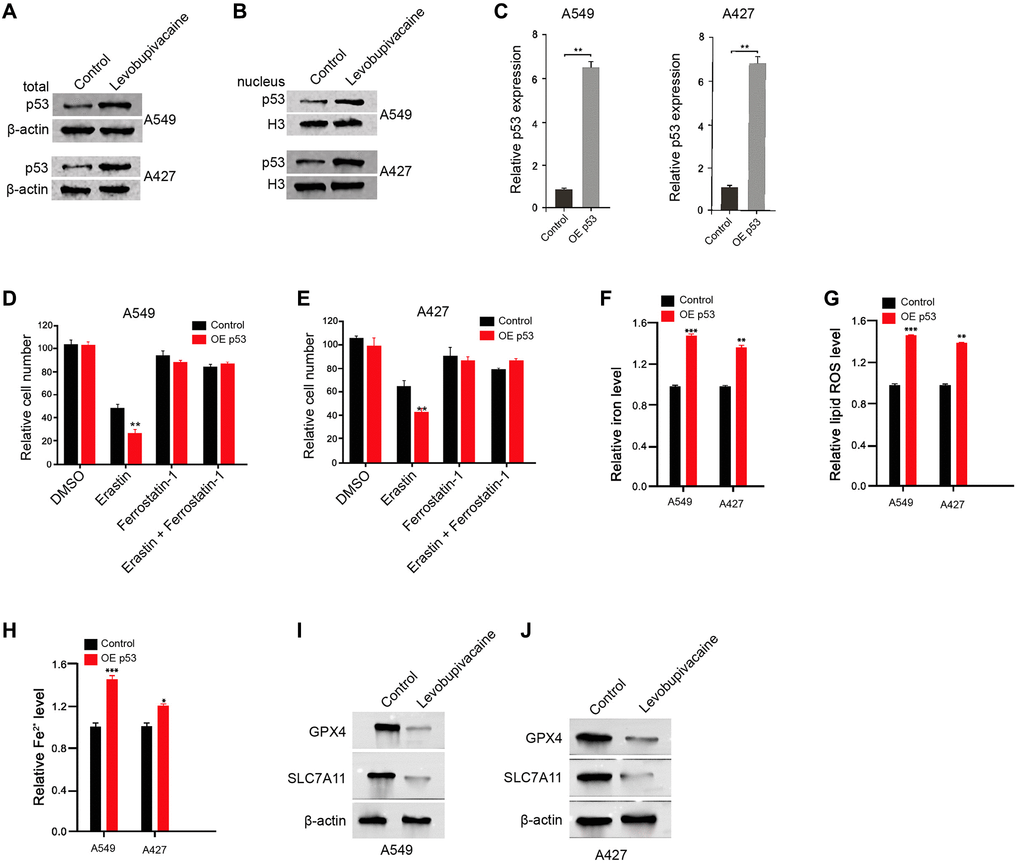
Figure 4. Levobupivacaine up-regulates and activates p53 in NSCLC cells. (A and B) The A549 and A427 cells were treated with levobupivacaine (2 mM) or equal volume saline. The total expression and nucleus accumulation of p53 was measured by Western blot analysis in the cells. (C) The A549 and A427 cells were treated with p53 overexpression vectors. (D and E) The A549 and A427 cells were co-treated with 5 mmol/L erastin or ferrostatin (1 mmol/L) and p53 overexpression vectors. The cell growth was analyzed by MTT assays. (F–J) The A549 and A427 cells were treated with p53 overexpression vectors or control vectors. (F) The levels of ROS were measure by flow cytometry analysis in the cells. (G and H) The levels of iron and Fe2+ were analyzed by Iron Assay Kit. (I and J) The expression of GPX4, SLC7A11, and β-actin was measured by Western blot analysis in the cells. Data are presented as mean ± SD. Statistic significant differences were indicated: *P < 0.05, **P < 0.01, ***P < 0.001.
Levobupivacaine induces ferroptosis by regulating p53 in NSCLC cells
Next, we tried to investigate whether levobupivacaine-induced ferroptosis by activating p53 in the NSCLC cells. The efficiency of p53 knockdown was confirmed in the cells (Figure 5A). The treatment of levobupivacaine promoted the erastin-induced inhibition of cell growth in the A549 and A427 cells, in which the depletion of p53 could rescue the phenotype (Figure 5B and 5C). Meanwhile, the treatment of levobupivacaine remarkably increased the levels of ROS, iron, Fe2+ in the A549 and A427 cells, but p53 knockdown reversed the effect (Figure 5D–5I). Similarly, the depletion of p53 was able to reverse levobupivacaine-inhibited GPX4 and SLC7A11 expression in the cells (Figure 5J). Taken together, these results suggest that levobupivacaine induces ferroptosis by regulating p53 in NSCLC cells.
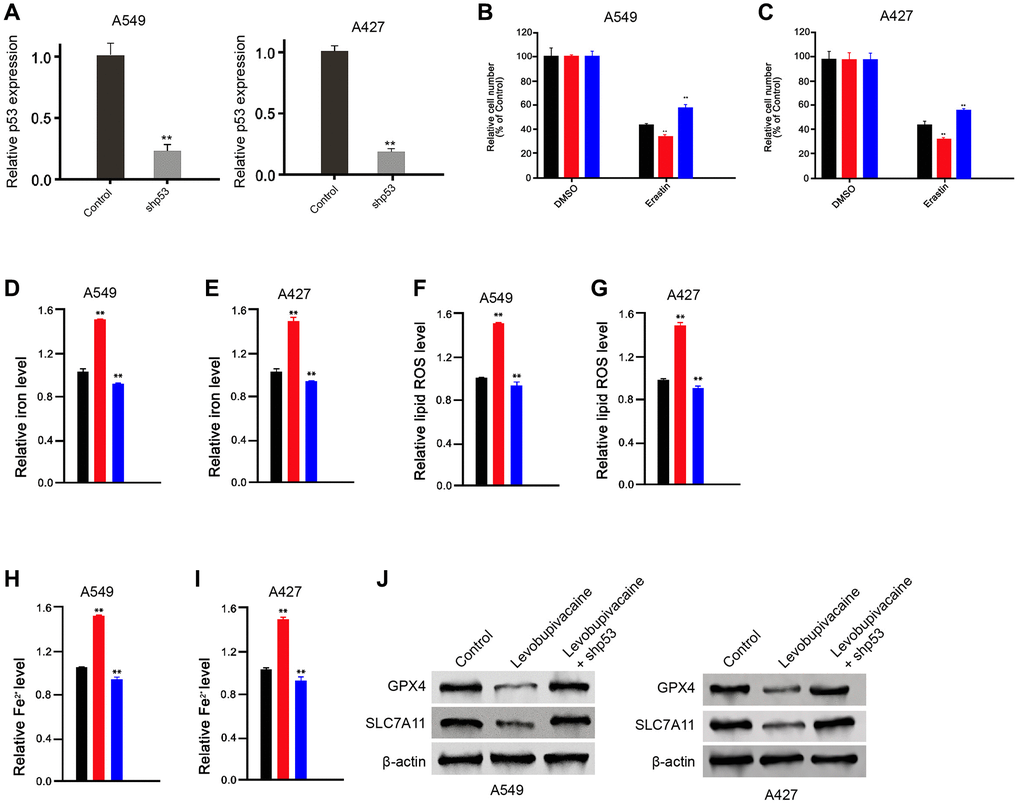
Figure 5. Levobupivacaine induces ferroptosis by regulating p53 in NSCLC cells. (A) The A549 and A427 cells were treated with p53 shRNA. The expression of p53 was measured by qPCR in the cells (B and C) The A549 and A427 cells were co-treated with 5 mmol/L erastin and 2 mM levobupivacaine, or co-treated with 5 mmol/L erastin, 2 mM levobupivacaine and p53 shRNA. The cell growth was analyzed by MTT assays. (D–J) The A549 and A427 cells were treated with levobupivacaine (2 mM) or co-treated with 2 mM levobupivacaine and p53 shRNA. (D and E) The levels of ROS were measure by flow cytometry analysis in the cells. (F–I) The levels of iron and Fe2+ were analyzed by Iron Assay Kit. (J) The expression of GPX4, SLC7A11, and β-actin was measured by Western blot analysis in the cells. Data are presented as mean ± SD. Statistic significant differences were indicated: *P < 0.01.
Levobupivacaine inhibits NSCLC progression by targeting p53
Next, we further detected the function of levobupivacaine/p53 axis in modulating of NSCLC progression. As expected, the depletion of p53 significantly reversed levobupivacaine-inhibited cell viability in the A549 and A427 cells (Figure 6A and 6B). The levobupivacaine-induced cell apoptosis was inhibited by p53 knockdown in the cells (Figure 6C and 6D). Together these data suggest that levobupivacaine inhibits NSCLC progression by targeting p53.
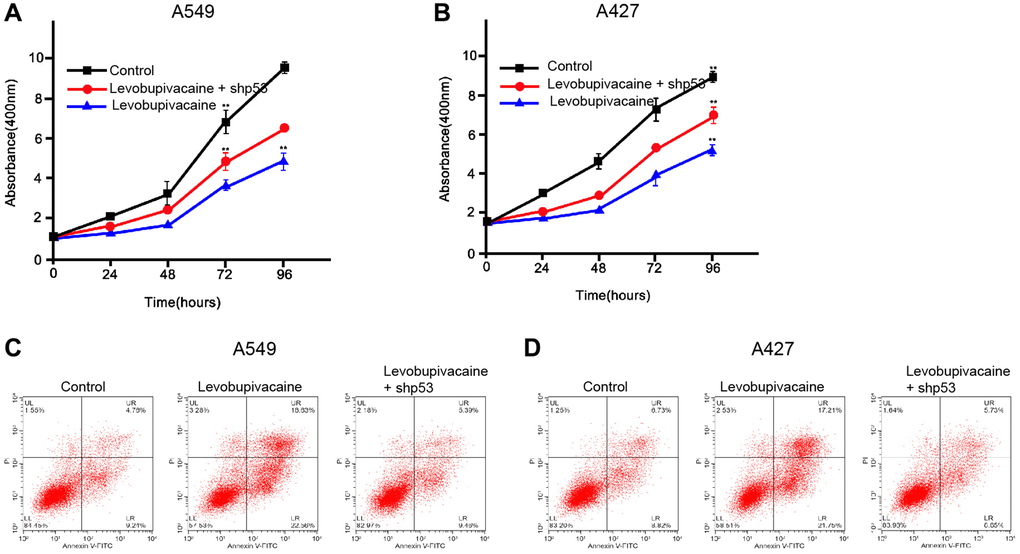
Figure 6. Levobupivacaine inhibits NSCLC progression by targeting p53. (A–D) The A549 and A427 cells were treated with levobupivacaine, or co-treated with levobupivacaine and p53 shRNA. (A and B) The cell viability was measured by MTT assays in the cells. (C and D) The cell apoptosis was measure by flow cytometry analysis in the cells. Data are presented as mean ± SD. Statistic significant differences were indicated: **P < 0.01.
Levobupivacaine attenuates the tumor growth of NSCLC in vivo
We further analyzed the impact of levobupivacaine on NSCLC development in vivo. For this purpose, we performed the tumorigenicity analysis in nude mice injected with A549 cells and the mice were treated with levobupivacaine. The treatment of levobupivacaine significantly repressed the tumor growth of A549 cells in vivo, as demonstrated by the tumor size (Figure 7A), tumor volume (Figure 7B), and tumor weight (Figure 7C). Besides, the expression of p53 was increased by the levobupivacaine treatment in the tumor tissues of the mice (Figure 7D). Together these indicate that levobupivacaine attenuates the tumor growth of NSCLC in vivo.
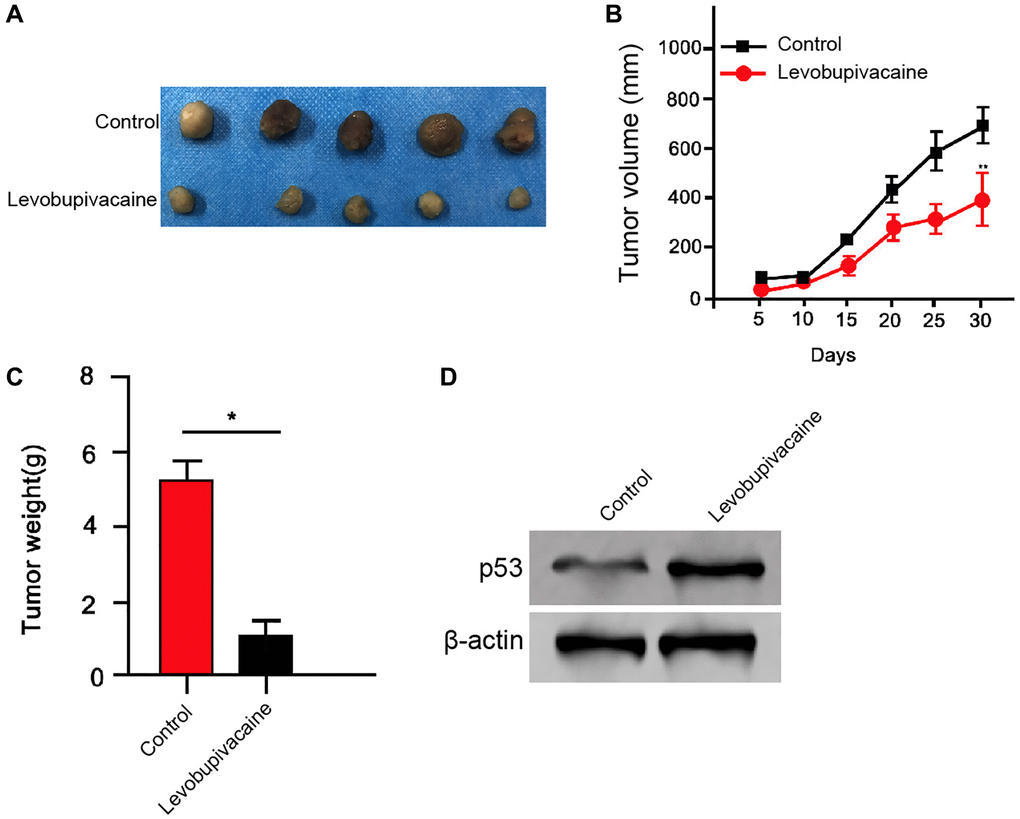
Figure 7. Levobupivacaine attenuates the tumor growth of NSCLC in vivo. (A–D) The effect of levobupivacaine on tumor growth of NSCLC cells in vivo was analyzed by nude mice tumorigenicity assay by injected with the A549 cells. The mice were treated with levobupivacaine (40 μmol/Kg) or equal volume saline. (A) Representative images of dissected tumors from nude mice were presented. (B) The average tumor volume was calculated and shown. (C) The average tumor weight was calculated and shown. (D) The expression of p53 and β-actin was measured by Western blot analysis in the tumor tissues of the mice. Data are presented as mean ± SD. Statistic significant differences were indicated: *P < 0.05, **P < 0.01.
Discussion
NSCLC is the prevalent lung cancer affecting more and more modern people and leading to severe mortality [20]. The local anesthetic levobupivacaine has presented potential anti-tumor activity in several cancer models [10, 11]. Nevertheless, the role of levobupivacaine in the development of NSCLC is still unreported. In this study, we firstly identified that levobupivacaine could induce ferroptosis and inhibit the progression of NSCLC by up-regulating and activating p53.
As a widely applied local anesthetic, levobupivacaine has shown potential anti-cancer activity in several cancer types. It has been reported that levobupivacaine induces an inhibitory effect on prostate cancer cell viability by modulating the Redox mechanism [10]. Levobupivacaine induces apoptosis and enhances proliferation by inhibiting the PI3K/Akt/mTOR signaling in breast cancer [11]. Levobupivacaine reduces the migration and proliferation in human melanoma and triple-negative breast cancer cells [21]. Levobupivacaine also inhibits the cell proliferation and invasion of breast cancer [22]. In this study, we identified that levobupivacaine inhibited proliferation, invasion, and migration, and induced apoptosis of NSCLC cells. Levobupivacaine was able to reduce the tumor growth of NSCLC in vivo. These data display a novel anti-tumor function of levobupivacaine in NSCLC progression, revealing important evidence of the function of the local anesthetic during NSCLC progresssion.
It has been recognized that the activation of ferroptosis is one potential therapeutic strategy [23]. Ferroptosis sensibility is influenced by several biological signaling, including epithelial-mesenchymal transition (EMT), metabolisms, DNA damage pathway, and loss of p53. Erastin can activate ferroptosis by inhibiting intracellular glutathione levels, reducing cystine import, and thereby leading to the redox imbalance [24–27]. Ferroptosis also presents fundamental roles during NSCLC development. Erastin-induced reactive oxygen species to up-regulate and activate p53 and promotes cytostatic and cytotoxic impacts in NSCLC cells [28]. Acetaminophen is able to sensitize erastin-activated ferroptosis via mediating Nrf2 signaling in NSCLC cells [29]. Erastin induces cisplatin-resistant and ferroptosis by inhibiting the Nrf2/xCT signaling in NSCLC cells [30]. NRF2 modulates ferroptosis in NSCLC cells by targeting FOCAD-FAK signaling [31]. Our investigation further demonstrated that levobupivacaine induced ferroptosis and inhibited progression of NSCLC by activating p53 in NSCLC cells. These data display an unreported correlation of levobupivacaine with ferroptosis and p53 in the development of NSCLC, identifying the new mechanism of levobupivacaine-induced anti-tumor activity. Meanwhile, the mechanism underlying the effect of levobupivacaine on p53 is not clear. It may be associated with transcriptional factors or ubiquitin enzymes, and needs further explore in future.
Conclusions
In conclusion, we discovered that the local anesthetic levobupivacaine inhibited the progression and induced ferroptosis of NSCLC by up-regulating p53. We present a novel mechanism underlying levobupivacaine-attenuated the development of NSCLC. Levobupivacaine may serve as the potential anti-cancer agent for NSCLC treatment.
Materials and Methods
Cell culture
The A549 and A427 cells were purchased from ATCC (USA). Cells were cultured at the incubator of 37 °C and 5% CO2, and incubated in RPMI-1640 (Gibco, USA) medium with FBS (10%, Gibco, USA), streptomycin (0.1 mg/mL, Gibco, USA), and penicillin (100 units/mL, Gibco, USA), and 10% fetal bovine serum (Gibco, USA). Levobupivacaine (Chirocaine, UK) was diluted by culture medium at the dose of 1 mM. The lentiviral plasmids carrying p53 shRNA, the corresponding control shRNA, the pcDNA3.1-p53 were synthesized (GenScript, China). Liposome 3000 (Invitrogen, USA) was used for cell transfection.
MTT assays
MTT assays were used to determine viability of A549 andA427 cells. Around 2 × 104 cells/well were seeded into 96-well plates and the measurement was performed every 24 hours from 0 hour to 72 hours. After indicated treatment, the cells were added with 10 μL, 5 mg/mL MTT agent for 4 hours and finished by 150 μL DMSO, followed by the observation using the ELISA browser (Bio-Tek EL 800, USA) at the absorbance of 570 nm.
Transwell assays
Transwell assays (Corning, USA) were applied to detect the migration and invasion of A549 and A427 cells. The 1 × 105 cells per well were seeded into upper chamber. The lower chamber was filled with 500 μL DMEM contained 10% FBS. The cells in the lower chamber were stained for 30 minutes using crystal violet after for 12 hours of the incubation. The migrated cell numbers were observed and counted using a microscope.
Colony formation assays
Approximately 1 × 103 A549 and A427 cells were seeded into 6-well plates and incubated for 2 weeks. After that, methanol was used to fix the cells and crystal violet (1%) was applied to dye the samples, followed by the observation, photographing, and calculation.
Wound healing assay
A549 and A427 cells were seeded into the 24-well plates at 3 × 105/well and cultured overnight to reach a full confluent as a monolayer. A 20 μl pipette tip was applied to slowly cut a straight line across the well. Then the well was washed by PBS 3 times and changed with the serum-free medium and continued to culture. The wound healing percentage was calculated.
Apoptosis analysis
Approximately 2 × 105 treated A549 and A427 cells were collected and subjected into apoptosis analysis based on Annexin V-FITC Apoptosis Detection Kit (CST, USA), in which the cells were dyed using Annexin V-FITC Apoptosis Detection Kit and detected using flow cytometry.
Ferroptosis analysis
The ferroptosis was analyzed as previous described [32]. The cells were co-treated with 5 mmol/L erastin or ferrostatin (1 mmol/L) and 1 mM levobupivacaine. MTT assays were used to analyze the cell viability. The reactive oxygen species levels were detected using flow cytometry analysis in the cells. The levels of Fe3+ and Fe2+ were analyzed by Iron Assay Kit (Abcam, USA) in the cells.
Cell-cycle analysis
Approximately 1 × 105 cells treated as indicated were plated into 6-well plates. Floating and adherent cells were fixed overnight in cold ethanol (4°C, 70% in PBS). RNaseA 1(00 μg/mL) was plated to the cells for 30 minutes at 37°C, followed by the PI staining (50 μg/mL, 30 minutes) and the flow cytometric analysis (Becton Dickinson, USA). About 10000 events were calculated per samples and distribution of cell cycles were calculated using Cell Quest software (Becton Dickinson, USA).
Western blot analysis
RIPA buffer (CST, USA) and BCA Protein Quantification Kit (Abbkine, USA) were used to isolated and quantified total proteins, respectively. Equal concentration of cell protein lysate was electrophoresed at the SDS-polyacrylamide gel (10%), followed by transferring to the PVDF membranes. The membranes were cultured with the corresponding primary antibodies (p53 (Abcam, USA), GPX4 (Abcam, USA), SLC7A11 (Abcam, USA), histone H3 (Abcam, USA) and β-actin (Abcam, USA)) at 4°C overnight, followed by the incubation using the corresponding secondary antibody (Abcam, USA). The samples were analyzed by chemiluminescence (ECL) and observed by Odyssey CLx Infrared Imaging System.
Analysis of tumorigenicity in nude mice
Balb/c nude mice (n = 5, 4-week-old, male) were applied to detect the impact of levobupivacaine on tumor growth. Mice subcutaneously injected with 1 × 107 A549 cells were treated with levobupivacaine (40 μmol/Kg) or equal volume saline. The tumor volume was remarked every 5 days and finished at 30 days after injection, followed by the analysis of volume (length × (width/2)2) and weight. Animal Ethics Committee of Jinan People’s Hospital approved the animal care and methods of this study.
Statistical analysis
Statistical analyses were conducted by utilizing the GraphPad prism 7 and were presented by mean ± SD. Two groups and multiple group differences were analyzed by unpaired Student's t-test and one-way ANOVA, respectively. P < 0.05 was expressed statistically significant.
Author Contributions
Minghua Meng, Meiling Huang, Jianhua Gu and Bin Ma contributed to the conception or design of the work. Jiming Wang, Weihua Ren, Jianhua Gu, Cuicui Liu and Shaomin Cui contributed to the acquisition, analysis, or interpretation of data for the work. Shuncheng He and Guangzhu Yang drafted the manuscript. Minghua Meng, Meiling Huang, Jianhua Gu, Bin Ma and Jinlan Xie critically revised the manuscript. All gave final approval and agree to be accountable for all aspects of work ensuring integrity and accuracy.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This research was supported by Jinan People’s Hospital Affiliated to Shandong First Medical University.
References
- 1. Hua Q, Mi B, Xu F, Wen J, Zhao L, Liu J, Huang G. Hypoxia-induced lncRNA-AC020978 promotes proliferation and glycolytic metabolism of non-small cell lung cancer by regulating PKM2/HIF-1α axis. Theranostics. 2020; 10:4762–78. https://doi.org/10.7150/thno.43839 [PubMed]
- 2. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018; 68:394–424. https://doi.org/10.3322/caac.21492 [PubMed]
- 3. Ettinger DS, Akerley W, Borghaei H, Chang AC, Cheney RT, Chirieac LR, D'Amico TA, Demmy TL, Ganti AK, Govindan R, Grannis FW
Jr , Horn L, Jahan TM, et al, and National Comprehensive Cancer Network (NCCN). Non-small cell lung cancer. J Natl Compr Canc Netw. 2012; 10:1236–71. https://doi.org/10.6004/jnccn.2012.0130 [PubMed] - 4. Nawaz K, Webster RM. The bladder cancer drug market. Nat Rev Drug Discov. 2016; 15:599–600. https://doi.org/10.1038/nrd.2016.141 [PubMed]
- 5. Kim C, Giaccone G. Precision oncology in non-small-cell lung cancer: opportunities and challenges. Nat Rev Clin Oncol. 2018; 15:348–49. https://doi.org/10.1038/s41571-018-0008-0 [PubMed]
- 6. Herbst RS, Morgensztern D, Boshoff C. The biology and management of non-small cell lung cancer. Nature. 2018; 553:446–54. https://doi.org/10.1038/nature25183 [PubMed]
- 7. Dan J, Gong X, Li D, Zhu G, Wang L, Li F. Inhibition of gastric cancer by local anesthetic bupivacaine through multiple mechanisms independent of sodium channel blockade. Biomed Pharmacother. 2018; 103:823–28. https://doi.org/10.1016/j.biopha.2018.04.106 [PubMed]
- 8. Xing W, Chen DT, Pan JH, Chen YH, Yan Y, Li Q, Xue RF, Yuan YF, Zeng WA. Lidocaine Induces Apoptosis and Suppresses Tumor Growth in Human Hepatocellular Carcinoma Cells In Vitro and in a Xenograft Model In Vivo. Anesthesiology. 2017; 126:868–81. https://doi.org/10.1097/ALN.0000000000001528 [PubMed]
- 9. Baydilek Y, Yurtlu BS, Hanci V, Ayoğlu H, Okyay RD, Kayhan GE, Tokgöz H, Mungan G, Ozkoçak I. The comparison of levobupivacaine in continuous or single dose spinal anesthesia for transurethral resection of prostate surgery. Braz J Anesthesiol. 2014; 64:89–97. https://doi.org/10.1016/j.bjane.2013.03.007 [PubMed]
- 10. Jose C, Hebert-Chatelain E, Dias Amoedo N, Roche E, Obre E, Lacombe D, Rezvani HR, Pourquier P, Nouette-Gaulain K, Rossignol R. Redox mechanism of levobupivacaine cytostatic effect on human prostate cancer cells. Redox Biol. 2018; 18:33–42. https://doi.org/10.1016/j.redox.2018.05.014 [PubMed]
- 11. Kwakye AK, Kampo S, Lv J, Ramzan MN, Richard SA, Falagán AA, Agudogo J, Atito-Narh E, Yan Q, Wen QP. Levobupivacaine inhibits proliferation and promotes apoptosis of breast cancer cells by suppressing the PI3K/Akt/mTOR signalling pathway. BMC Res Notes. 2020; 13:386. https://doi.org/10.1186/s13104-020-05191-2 [PubMed]
- 12. Battaglia AM, Chirillo R, Aversa I, Sacco A, Costanzo F, Biamonte F. Ferroptosis and Cancer: Mitochondria Meet the "Iron Maiden" Cell Death. Cells. 2020; 9:1505. https://doi.org/10.3390/cells9061505 [PubMed]
- 13. Dolma S, Lessnick SL, Hahn WC, Stockwell BR. Identification of genotype-selective antitumor agents using synthetic lethal chemical screening in engineered human tumor cells. Cancer Cell. 2003; 3:285–96. https://doi.org/10.1016/s1535-6108(03)00050-3 [PubMed]
- 14. Chen X, Li J, Kang R, Klionsky DJ, Tang D. Ferroptosis: machinery and regulation. Autophagy. 2020. [Epub ahead of print]. https://doi.org/10.1080/15548627.2020.1810918 [PubMed]
- 15. Yang WS, Stockwell BR. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells. Chem Biol. 2008; 15:234–45. https://doi.org/10.1016/j.chembiol.2008.02.010 [PubMed]
- 16. Yee PP, Wei Y, Kim SY, Lu T, Chih SY, Lawson C, Tang M, Liu Z, Anderson B, Thamburaj K, Young MM, Aregawi DG, Glantz MJ, et al. Neutrophil-induced ferroptosis promotes tumor necrosis in glioblastoma progression. Nat Commun. 2020; 11:5424. https://doi.org/10.1038/s41467-020-19193-y [PubMed]
- 17. Jiang M, Qiao M, Zhao C, Deng J, Li X, Zhou C. Targeting ferroptosis for cancer therapy: exploring novel strategies from its mechanisms and role in cancers. Transl Lung Cancer Res. 2020; 9:1569–84. https://doi.org/10.21037/tlcr-20-341 [PubMed]
- 18. Lou JS, Zhao LP, Huang ZH, Chen XY, Xu JT, Tai WC, Tsim KWK, Chen YT, Xie T. Ginkgetin derived from Ginkgo biloba leaves enhances the therapeutic effect of cisplatin via ferroptosis-mediated disruption of the Nrf2/HO-1 axis in EGFR wild-type non-small-cell lung cancer. Phytomedicine. 2021; 80:153370. https://doi.org/10.1016/j.phymed.2020.153370 [PubMed]
- 19. Gnanapradeepan K, Basu S, Barnoud T, Budina-Kolomets A, Kung CP, Murphy ME. The p53 Tumor Suppressor in the Control of Metabolism and Ferroptosis. Front Endocrinol (Lausanne). 2018; 9:124. https://doi.org/10.3389/fendo.2018.00124 [PubMed]
- 20. Valentino F, Borra G, Allione P, Rossi L. Emerging targets in advanced non-small-cell lung cancer. Future Oncol. 2018; 14:61–72. https://doi.org/10.2217/fon-2018-0099 [PubMed]
- 21. Castelli V, Piroli A, Marinangeli F, d'Angelo M, Benedetti E, Ippoliti R, Zis P, Varrassi G, Giordano A, Paladini A, Cimini A. Local anesthetics counteract cell proliferation and migration of human triple-negative breast cancer and melanoma cells. J Cell Physiol. 2020; 235:3474–84. https://doi.org/10.1002/jcp.29236 [PubMed]
- 22. Li R, Xiao C, Liu H, Huang Y, Dilger JP, Lin J. Effects of local anesthetics on breast cancer cell viability and migration. BMC Cancer. 2018; 18:666. https://doi.org/10.1186/s12885-018-4576-2 [PubMed]
- 23. Liang C, Zhang X, Yang M, Dong X. Recent Progress in Ferroptosis Inducers for Cancer Therapy. Adv Mater. 2019; 31:e1904197. https://doi.org/10.1002/adma.201904197 [PubMed]
- 24. Xie Y, Zhu S, Song X, Sun X, Fan Y, Liu J, Zhong M, Yuan H, Zhang L, Billiar TR, Lotze MT, Zeh HJ
3rd , Kang R, et al. The Tumor Suppressor p53 Limits Ferroptosis by Blocking DPP4 Activity. Cell Rep. 2017; 20:1692–704. https://doi.org/10.1016/j.celrep.2017.07.055 [PubMed] - 25. Chen PH, Wu J, Ding CC, Lin CC, Pan S, Bossa N, Xu Y, Yang WH, Mathey-Prevot B, Chi JT. Kinome screen of ferroptosis reveals a novel role of ATM in regulating iron metabolism. Cell Death Differ. 2020; 27:1008–22. https://doi.org/10.1038/s41418-019-0393-7 [PubMed]
- 26. Doll S, Proneth B, Tyurina YY, Panzilius E, Kobayashi S, Ingold I, Irmler M, Beckers J, Aichler M, Walch A, Prokisch H, Trümbach D, Mao G, et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat Chem Biol. 2017; 13:91–98. https://doi.org/10.1038/nchembio.2239 [PubMed]
- 27. Chen PH, Smith TJ, Wu J, Siesser PF, Bisnett BJ, Khan F, Hogue M, Soderblom E, Tang F, Marks JR, Major MB, Swarts BM, Boyce M, Chi JT. Glycosylation of KEAP1 links nutrient sensing to redox stress signaling. EMBO J. 2017; 36:2233–50. https://doi.org/10.15252/embj.201696113 [PubMed]
- 28. Huang C, Yang M, Deng J, Li P, Su W, Jiang R. Upregulation and activation of p53 by erastin-induced reactive oxygen species contribute to cytotoxic and cytostatic effects in A549 lung cancer cells. Oncol Rep. 2018; 40:2363–70. https://doi.org/10.3892/or.2018.6585 [PubMed]
- 29. Gai C, Yu M, Li Z, Wang Y, Ding D, Zheng J, Lv S, Zhang W, Li W. Acetaminophen sensitizing erastin-induced ferroptosis via modulation of Nrf2/heme oxygenase-1 signaling pathway in non-small-cell lung cancer. J Cell Physiol. 2020; 235:3329–39. https://doi.org/10.1002/jcp.29221 [PubMed]
- 30. Li Y, Yan H, Xu X, Liu H, Wu C, Zhao L. Erastin/sorafenib induces cisplatin-resistant non-small cell lung cancer cell ferroptosis through inhibition of the Nrf2/xCT pathway. Oncol Lett. 2020; 19:323–33. https://doi.org/10.3892/ol.2019.11066 [PubMed]
- 31. Liu P, Wu D, Duan J, Xiao H, Zhou Y, Zhao L, Feng Y. NRF2 regulates the sensitivity of human NSCLC cells to cystine deprivation-induced ferroptosis via FOCAD-FAK signaling pathway. Redox Biol. 2020; 37:101702. https://doi.org/10.1016/j.redox.2020.101702 [PubMed]
- 32. Mao C, Wang X, Liu Y, Wang M, Yan B, Jiang Y, Shi Y, Shen Y, Liu X, Lai W, Yang R, Xiao D, Cheng Y, et al. A G3BP1-Interacting lncRNA Promotes Ferroptosis and Apoptosis in Cancer via Nuclear Sequestration of p53. Cancer Res. 2018; 78:3484–96. https://doi.org/10.1158/0008-5472.CAN-17-3454 [PubMed]