



Introduction
Sestrins were identified two decades ago as stress-responsive proteins that play an important role in regulating cellular homeostasis. Vertebrate genomes showcase three Sestrin genes (SESN1-3), while invertebrates feature just one [1–3]. Numerous stressors, ranging from hypoxia and oxidative stress to DNA damage and nutrient deprivation, induce Sestrin expression in mammalian cells. The orchestration behind this expression involves several transcription factors, notably p53, FOXO, ATF4 and NRF2 [4–6]. Highlighting evolutionary conservation [7], the same signalling pathways trigger the activation of dSesn in D. melanogaster [8]. Consequently, Sestrins play pivotal roles in the regulation of cellular viability under various stress conditions, such as hypoxia, oxidative stress, DNA damage and glucose deprivation [2, 9–13].
Earlier research from our team established Sestrins as antioxidant proteins that play a critical role in inhibiting the mechanistic target of rapamycin complex 1 (mTORC1) kinase [9, 14–16]. mTORC1 is an intricate environmental sensor that integrates signals from nutrients, growth factors and stresses to regulate cell fate decisions. mTORC1 functions as a central regulator of biosynthesis and cell growth, while also suppressing macroautophagy (herein in the text – autophagy) [17]. Autophagy is the process of encapsulating intracellular components into autophagosomal vesicles, followed by the degradation of their contents in lysosomes. Autophagy is essential for nutrient supply and cell repair under stressful conditions. While autophagy typically supports cell survival under stress, it can also trigger autophagy-dependent cell death [18].
Remarkably, mTORC1 plays a key role in lifespan and aging regulation across various species. Application of specific mTORC1 inhibitors, like rapamycin, has been shown to enhance lifespan in different organisms from yeast to mice [19–23]. Similarly, caloric restriction (CR), a well-documented longevity enhancer across many species, also represses mTORC1 activity, further cementing the role of this kinase in aging control [24–26]. Nutrient and energy availability signals are transmitted to mTORC1 through the evolutionarily conserved insulin/IGF1 signaling pathway in metazoans [27]. In addition to mTORC1, the insulin/IGF1 signalling pathway inhibits the forkhead box O (FoxO) transcription factors via Akt-mediated phosphorylation and nuclear exclusion [28], and FoxOs regulate adaptation to starvation conditions in metazoans [29, 30]. In contrast, mTORC1 is activated by Akt in response to insulin/IGF-1 signaling and promotes anabolic processes while inhibiting catabolic pathways [31].
Branched-chain amino acids (BCAA), especially leucine, play a critical role in the regulation of mTORC1, primarily via activation of the GATOR2 protein complex. GATOR2 is an inhibitor of the GATOR1 protein complex, which suppresses the activity of RagA/B GTPases, thereby preventing mTORC1 translocation to lysosomes and its activation [17]. Sestrins inhibit mTORC1 through the interaction with GATOR2. Leucine binding to Sestrins induces conformational changes that weaken the interaction between Sestrins and GATOR2, leading to mTORC1 activation [32, 33].
Being regulators of stress response and metabolism, broader implications of Sestrins in aging control cannot be understated. In C. elegans, variations in sesn-1 expression levels have measurable effects on lifespan and physiological functions [34, 35]. Similarly, in D. melanogaster, alterations in dSesn levels contribute to the development of age-associated disorders such as muscle degeneration and cardiac arrhythmia [8]. Many of Sestrin’s effects might be attributed to its role in activating autophagy through the mTORC1-mediated mechanism. Recent findings indicate that dSesn plays a key role in extending lifespan in flies subjected to BCAA restriction [36]. Stem cell functionality is maintained through autophagy [37] and the impact of Sestrins on various facets of stem cell biology, encompassing both stemness and differentiation, is also being recognized [38].
In an effort to elucidate overarching role of Sestrins in lifespan modulation during caloric deprivation (CD), we utilized a C. elegans model in which sesn-1 was inactivated via gene deletion [35]. Existing research has underscored the remarkable lifespan extension in nematodes upon CD [39, 40]. Our studies aimed to elucidate Sestrin’s role in lifespan regulation under CD and to examine how sesn-1 deficient worms respond to stress. Based on our studies in C. elegans, we demonstrate that the relationship between sesn-1 and the GATOR–TORC1–autophagy axis is highly conserved across eukaryotes [7]. We investigated the involvement of sesn-1 in signalling pathways that link mTORC1 activity, autophagy and increased lifespan during CD in C. elegans, such as those regulated by let-363 (an ortholog of the mammalian mTOR gene) [41], daf-2 (an ortholog of the mammalian IGFR1 gene) [42] and daf-16 (an ortholog of the mammalian FOXO genes) [42]. We also evaluated the potential involvement of sesn-1 in lifespan extension in nematodes carrying a deletion in the eat-2 gene [43] that is required for proper pharyngeal function. Animals carrying this mutation experience continuous, moderate caloric restriction throughout development and adulthood. Our findings confirm the critical role of sesn-1 in lifespan extension mediated by mTORC1 inhibition and autophagy activation in response to CD.
Results
Sesn-1 modulates lifespan extension in C. elegans
To understand the role of sesn-1, the nematode ortholog of the Sestrin genes, in the control of aging and lifespan, we determined its impact on lifespan regulation under CD conditions. Using established protocols [40], nematodes were cultured on agar plates with (control) or without bacteria to assess lifespan. As previously reported [34], sesn1-deletion mutant strain, sesn-1(ok3157), exhibited a marginally reduced lifespan compared to their wild-type (WT) counterparts under normal conditions. This suggests that sesn-1 facilitates nematode homeostasis in food-abundant conditions. However, in our observations, the lifespan of the sesn-1(ok3157) animals did not differ from that of the WT counterparts under ad libitum conditions (Figure 1 and Table 1). Therefore, we decided to examine the role of sesn-1 in the regulation of lifespan in response to CD. According to our data, CD augmented the lifespan of WT animals by 40.2%. In contrast, the sesn-1(ok3157) worms experienced a mere 6.2% increase in lifespan, underscoring the pivotal role of sesn-1 in lifespan extension following CD exposure (Figure 1 and Table 1). The same data were observed in another sesn-1 deficient strain, sesn-1(ie24589) (IE24589 strain with MOS-1 transposon insertion in 3 exon) (Supplementary Figure 1).
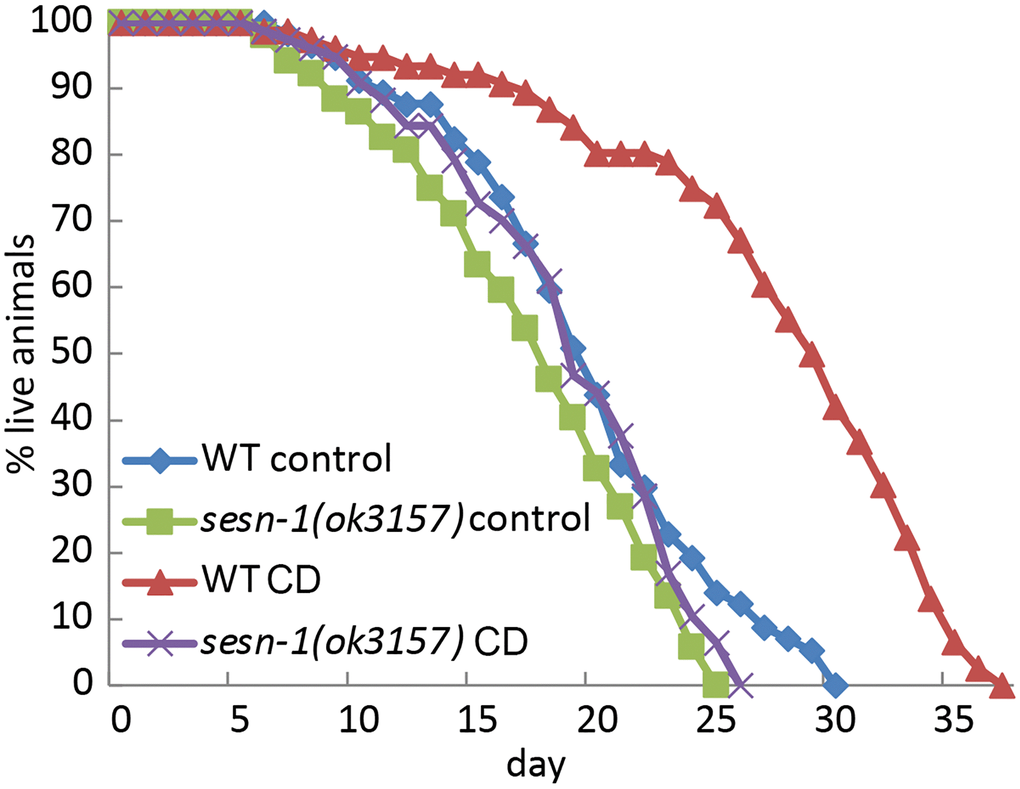
Figure 1. C. elegans lifespan extension under CD is modulated by sesn-1. The lifespan of WT and sesn-1(ok3157) nematodes was assessed after plating on control or axenic media.
Table 1. Lifespan extension means analysis for control WT and sesn-1(ok3157) nematodes under caloric deprivation, with RNAi expression against npp-18 and Y32H12A.8.
| Strain | RNAi | Control mean lifespan ± SEM, days | n | Starvation mean lifespan ± SEM, days | n | Effect vs. control % | p-value vs. control |
| WT | EV | 18,1 ± 0,50 | 46 | 24,5 ± 0,61 | 57 | + 40,2% | <0,0001 |
| sesn-1(ok3157) | EV | 17,6 ± 0,52 | 49 | 18,8 ± 0,6 | 57 | + 6,19% | 0,8242 |
| WT | npp-18 | 17,5 ± 0,47 | 52 | 18,3 ± 0,58 | 49 | + 4,38% | >0,9999 |
| sesn-1(ok3157) | npp-18 | 16,7 ± 0,44 | 51 | 17,3 ± 0,54 | 52 | + 3,95% | >0,9999 |
| WT | Y32H12A.8 | 17,1 ± 0,59 | 50 | 17,8 ± 0,71 | 46 | + 3,5% | >0,9999 |
| sesn-1(ok3157) | Y32H12A.8 | 16,5 ± 0,53 | 50 | 16,5 ± 0,45 | 44 | + 0,12% | >0,9999 |
Sesn-1 protects nematodes from multiple stresses
Given the link between stress response and lifespan modulation, where proteins responsive to stress curb accumulation of age-linked damage, we investigated how sesn-1 influences stress tolerance. Exposing nematodes to various stressors like oxidizing agents (paraquat and hydrogen peroxide) and axenic culture medium (M9) revealed stark differences between WT and sesn-1(ok3157) or sesn-1(ie24589) animals. When WT first larval stage (L1) nematodes were placed in M9 medium, they outlived their sesn-1(ok3157) or sesn-1(ie24589) congeners, showing reduced resistance of sesn-1 mutants to nutrient restriction (Figure 2A). Furthermore, oxidative stress induced by paraquat and H2O2 dramatically accelerated the death of sesn-1(ok3157) and sesn-1(ie24589) worms compared to their WT counterparts (Figure 2B, 2C). These collective findings emphasize the role of sesn-1 in bolstering stress resilience in nematodes, likely through mechanisms analogous to those that promote lifespan extension during CD.
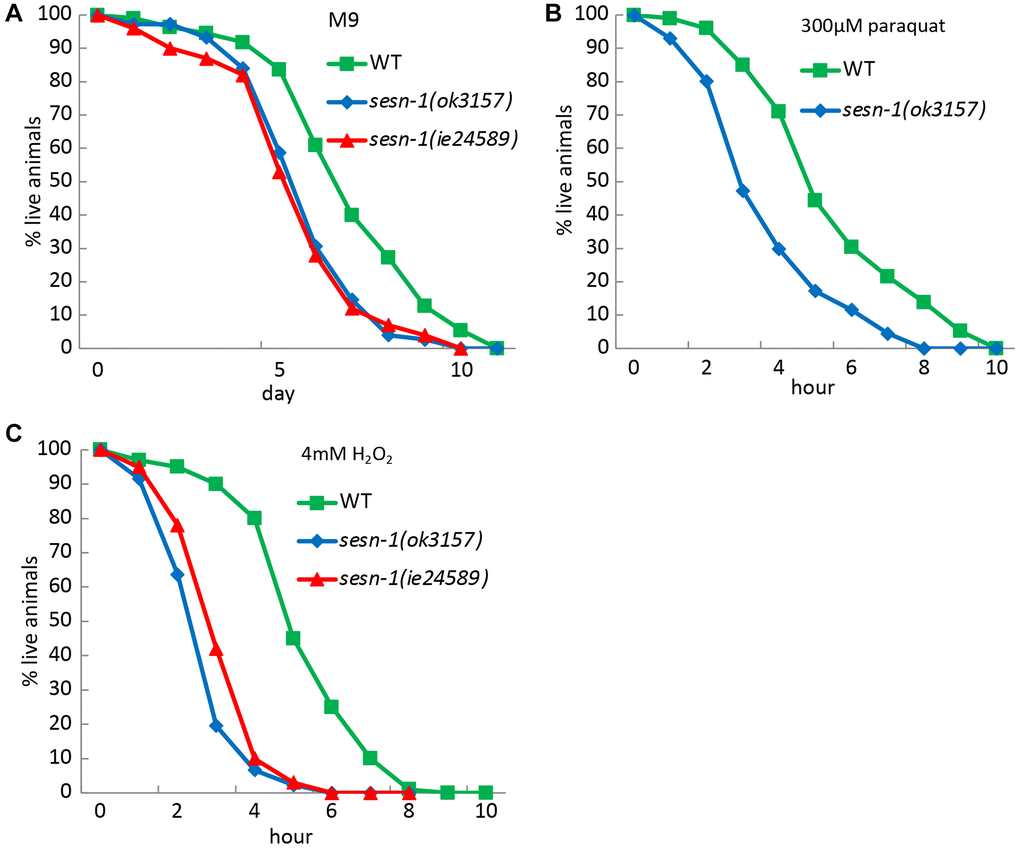
Figure 2. Role of Sesn-1 in stress resistance. The viability of WT and sesn-1(ok3157) C. elegans was studied under various stress conditions. (A) In axenic M9 medium (n = 783 for WT, n = 437 for sesn-1(ok3157), n = 511 for sesn-1(ie24589), the mean survival rates for sesn-1(ok3157) and sesn-1(ie24589) were 6.13 ± 0.4 and 6.16 ± 0.3 days, respectively, compared to 7.9 ± 0.5 days for WT. The difference between sesn-1(ok3157) and sesn-1(ie24589) was not significant (P > 0.05), while both mutants showed significantly lower survival than WT (P < 0.001). (B) In the presence of 300 μM paraquat (n = 1103 for WT, n = 876 for sesn-1(ok3157), n = 882 for sesn-1(ie24589)), the mean survival rates were 4.1 ± 1.2 and 4.0 ± 0.9 hours for sesn-1(ok3157) and sesn-1(ie24589), respectively, compared to 5.8 ± 1.2 hours for WT. Again, the difference between the two sesn-1 mutants was not significant (P > 0.05), while both were significantly more sensitive than WT (P < 0.001). (C) In the presence of 4 mM H2O2 (n = 1301 for WT, n = 930 for sesn-1(ok3157), n = 827 for sesn-1(ie24589)), the mean survival rates were 3.3 ± 0.5 and 3.8 ± 0.6 hours for sesn-1(ok3157) and sesn-1(ie24589), respectively, compared to 6.4 ± 0.8 hours for WT. The difference between sesn-1(ok3157) and sesn-1(ie24589) was not statistically significant (P > 0.05), while both mutants showed significantly reduced survival compared to WT (P < 0.001). Data are presented as mean ± S.E.M.
Sesn-1: an essential component for autophagy activation
Autophagy, a mechanism that promotes lifespan extension during nutrient scarcity and augments stress resistance [44, 45], may be activated by sesn-1. To test this hypothesis, we utilized a C. elegans strain adls2122 that expresses a GFP-tagged LGG-1 fusion protein (LGG-1::GFP), where LGG-1 is the nematode ortholog of the mammalian autophagy marker LC3. Autophagosomes incorporating LGG-1::GFP form discrete GFP-positive vesicles, which can be readily visualized by fluorescence microscopy. [46]. In WT third larval stage (L3) nematodes subjected to CD, we observed a pronounced accumulation of LGG-1::GFP-labeled autophagosomes within the seam cells. In contrast, nematodes with sesn-1 silenced by RNAi (sesn-1(RNAi)) exhibited only a modest increase in LGG-1::GFP-labeled autophagosomes, emphasizing sesn-1’s crucial role in autophagy initiation during starvation. Under control conditions, WT animals exhibited an average of 0.38 autophagosomes per cell compared to just 0.14 autophagosomes per cell in sesn-1 mutants—a difference of more than two-fold, which was statistically significant (p = 0.02194) (Figure 3A). In mammals, Sestrin-dependent autophagy activation is mediated by mTORC1 inhibition [6]. To ascertain sesn-1’s role in autophagy during nematode starvation, we subjected both WT and sesn-1(RNAi) worms to CD and evaluated mTORC1 activity and autophagy levels using immunoblotting. While the control group exhibited reduced ribosomal protein S6 phosphorylation post-CD exposure — an mTORC1-inhibiting event — this reduction was not observed in sesn-1(RNAi) animals (Figure 3B). Evaluation of autophagy by comparing LGG-1::GFP with its pro-autophagic, phosphatidylethanolamine-conjugated form (LGG-1::GFP-PE) revealed that CD prompted substantial accumulation of LGG-1::GFP-PE in WT worms. Yet, sesn-1(RNAi) worms exhibited only a minor increase in this autophagosome marker post-starvation, again highlighting sesn-1’s indispensable role in autophagy modulation under CD in C. elegans (Figure 3B). In a bid to elucidate sesn-1 contribution to autophagosome formation, we evaluated LGG-1::GFP-PE formation intensity in worms pre-treated with 200 mM chloroquine for 24 hours [47, 48]. Under acute starvation, WT worms manifested pronounced LGG-1::GFP-PE accumulation, but this was notably suppressed in sesn-1(RNAi) worms, signifying sesn-1’s necessity for appropriate autophagy activation (Supplementary Figure 2).
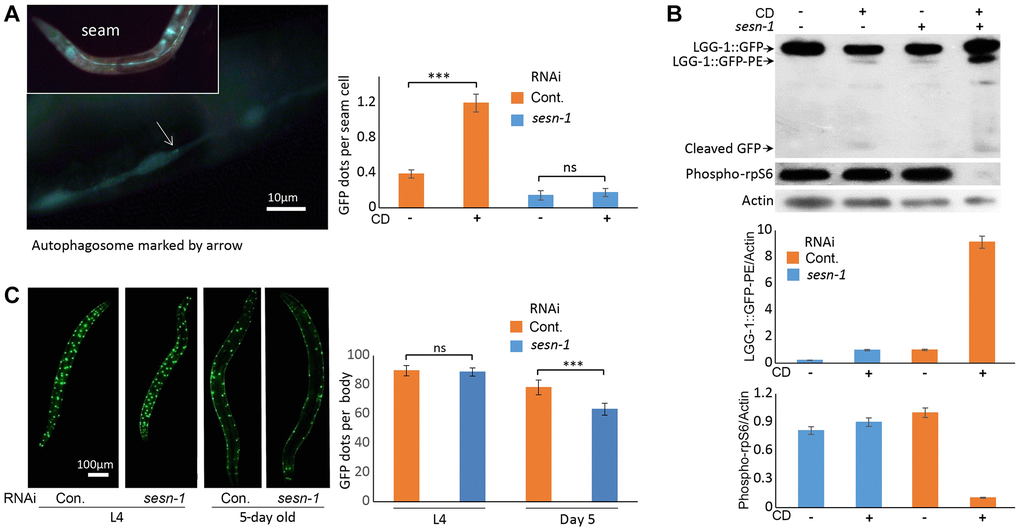
Figure 3. Autophagy activation by sesn-1 under CD correlates with reduced muscle degeneration. (A) Autophagosome accumulation in seam cells. Both control adIs2122 (DA2123 strain) and adIs2122; sesn-1(RNAi) nematodes, expressing a GFP-tagged LGG-1 fusion protein during L3, were exposed to axenic medium. Autophagosome counts per seam cell were analyzed under control conditions (n = 137 for adIs2122, n = 56 for adIs2122; sesn-1(RNAi)) and starvation conditions (n = 117 for adIs2122, n = 80 for adIs2122; sesn-1(RNAi)). “ns” and “***” indicate P-values > 0.05 and < 0.001, respectively. All bar graphs are presented as mean ± S.E.M. (B) Immunoblot and densitometric analyses showing relative levels of GFP::LGG-1, its phosphatidylethanolamine-conjugated form (LGG-1::GFP-PE), and the phosphorylated form of ribosomal protein S6 (phospho-rpS6) in adIs2122 (DA2123 strain) and adIs2122; sesn-1(RNAi) worms. All bar graphs represent blot intensity normalized to actin. (C) Whole-body images of nematodes expressing a myo-3p::GFP NLS-tagged fusion protein in body wall muscle nuclei in the ccIs4251 (PD4251 strain). Nuclear counts were performed at L4 (n = 24 for ccIs4251 and n = 23 for ccIs4251; sesn-1(RNAi)) and at 5 days of adulthood (n = 20 for both groups). “ns” and “***” indicate P-values > 0.05 and < 0.001, respectively. All bar graphs are presented as mean ± S.E.M.
Role of Sesn-1 role in sustaining muscle integrity
Previous studies in D. melanogaster and mice have linked Sestrins to preservation of muscle function, primarily through their role in mitigating oxidative stress-induced damage [8, 38, 49]. To investigate sesn-1’s potential role in preserving muscle density in C. elegans, we utilized a nematode strain (ccIs4251 I, e1282 IV) expressing myo-3p::GFP NLS-tagged fusion protein, which labels myocyte nuclei. Assessment of muscle density across various developmental stages, particularly the fourth larval stage (L4) and 5-day-old adult stage, in WT and sesn-1(RNAi) animals revealed that muscle density was consistent in the L4 animals across the groups. However, a pronounced reduction in myocyte count was observed in 5-day-old sesn-1(RNAi) adult worms, highlighting the essential role of sesn-1 in maintaining adult muscle function (Figure 3C).
Sesn-1 facilitates lifespan extension through GATOR2
Previous studies in mammalian cells have shown that Sestrins suppress mTORC1 activity by inhibiting GATOR2 [32, 50]. The Sestrin-GATOR-mTORC1 signalling pathway is known to be conserved in eukaryotes [7], so we analysed the potential involvement of GATOR2 in the sesn-1-modulated lifespan extension. We proposed that if sesn-1’s effects on the lifespan extension are GATOR2-dependent, sesn-1 would not significantly influence lifespan in GATOR2-deficient worms under CD. To test this, we used RNAi to silence the genes encoding the major components of GATOR2: npp-18 and Y32H12A.8, the orthologs of the mammalian SEH1L and WDR24 genes, respectively, in WT and sesn-1(ok3157) nematodes (Figure 4A, 4B) and measured lifespan increase during CD. While sesn-1 facilitated lifespan extension during CD in control worms, its effects were notably diminished when npp-18 and Y32H12A.8 were suppressed (Figure 4A, 4B and Table 1). Further studies examining sesn-1’s role in autophagy regulation in worms with suppressed GATOR2, using immunoblotting, revealed intriguing findings. In the absence of sesn-1, worms still exhibited modest autophagy; however, simultaneous RNAi-mediated suppression of sesn-1 and npp-18 significantly reduced autophagy. This suggests that sesn-1 likely operates via npp-18 (Figure 5).
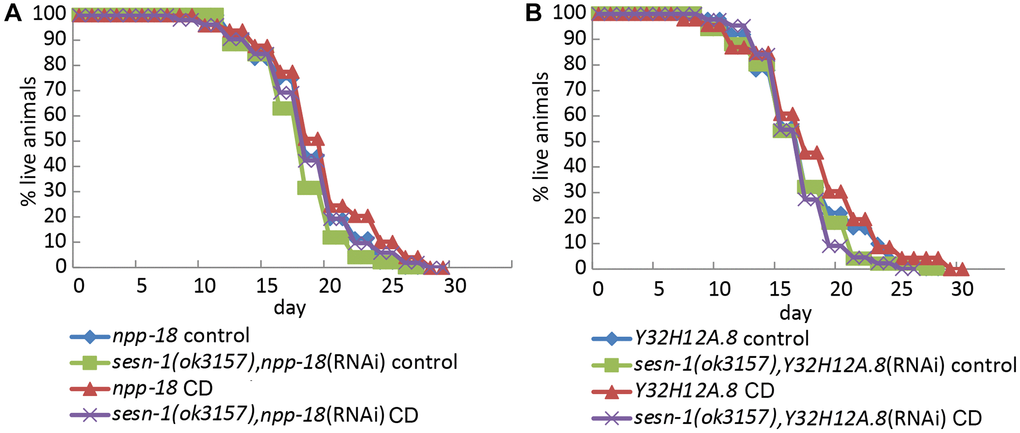
Figure 4. Sesn-1 modulates longevity under CD via GATOR2. (A) Lifespan of WT and sesn-1(ok3157) worms subjected to npp-18 RNAi knockdown in ad libitum and axenic media. (B) Lifespan of WT and sesn-1(ok3157) nematodes subjected to Y32H12A.8 RNAi knockdown in ad libitum and axenic media.
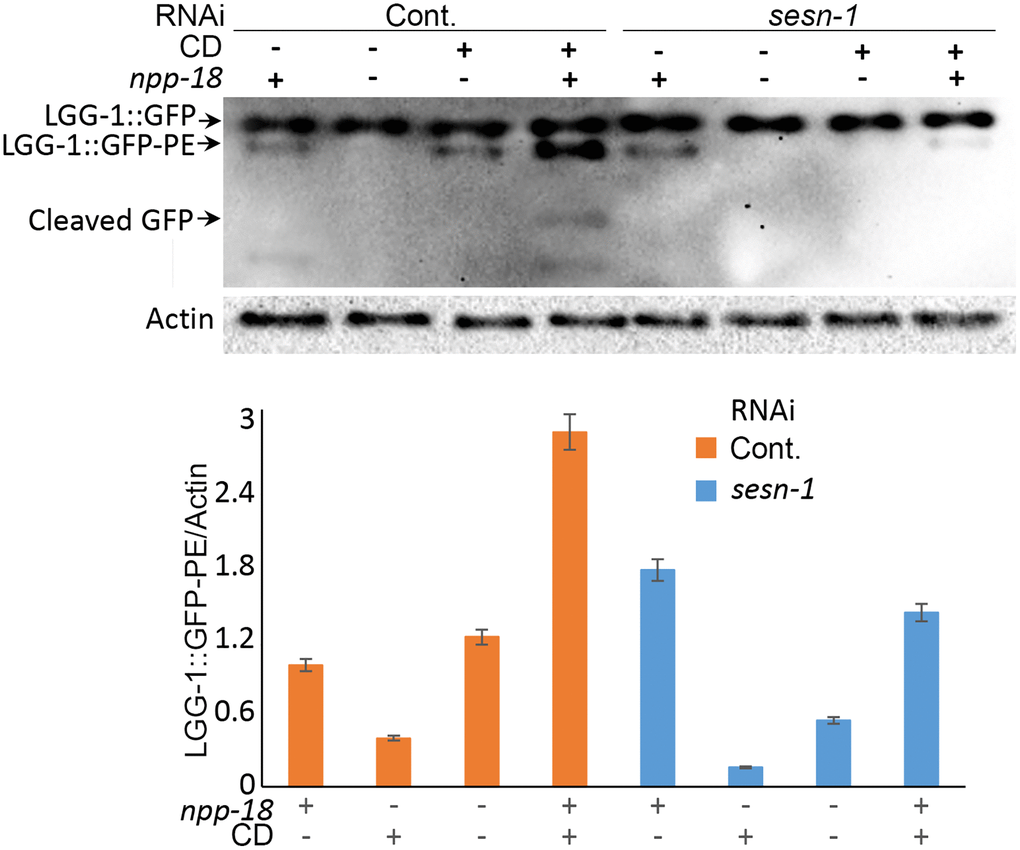
Figure 5. Autophagy intensity in C. elegans is influenced by sesn-1 and npp-18, a component of the GATOR2 complex. Control adIs2122 and adIs2122; sesn-1(RNAi) nematodes expressing GFP::LGG-1, with or without simultaneous npp-18 knockdown by RNAi, were subjected to starvation. The relative levels of GFP::LGG-1 conjugation to autophagosomes were measured by immunoblot densitometry. All bar graphs represent blot intensity normalized to actin.
Sesn-1 supports lifespan extension via FOXO and in eat-2 mutants
Sesn-1 may be involved in the signaling pathways known to be controlled by CD in C. elegans, such as those regulated by let-363(TOR) [41, 51], daf-2(IGF1R) [42] and daf-16(FOXO) [42]. We also tested the possible involvement of sesn-1 in lifespan extension in nematodes with a deletion in the eat-2 (ad1116) gene [43]. We used nematode strains with knockouts of eat-2(ad1116), daf-2(e1370) and daf-16(mu86) either with or without sesn-1 silencing by RNAi. The contribution of sesn-1 to the FOXO pathway and lifespan extension in eat-2(ad1116) nematodes was appreciable but did not reach statistical significance (Figure 6A–6D and Table 2). Without daf-16 and sesn-1, the lifespan of starved worms is reduced by 9.5% (Figure 6C), whereas the presence of sesn-1 increases lifespan of daf-16(mu86) worms by 7.5% under CD conditions. Survival of the eat-2 worms under CD requires sufficient autophagic activity. In starved nematodes lacking both eat-2 and sesn-1, lifespan increased by 3.6%, whereas the presence of sesn-1 extended the lifespan of eat-2(ad1116) mutants by 11% (Figure 6D and Table 2).
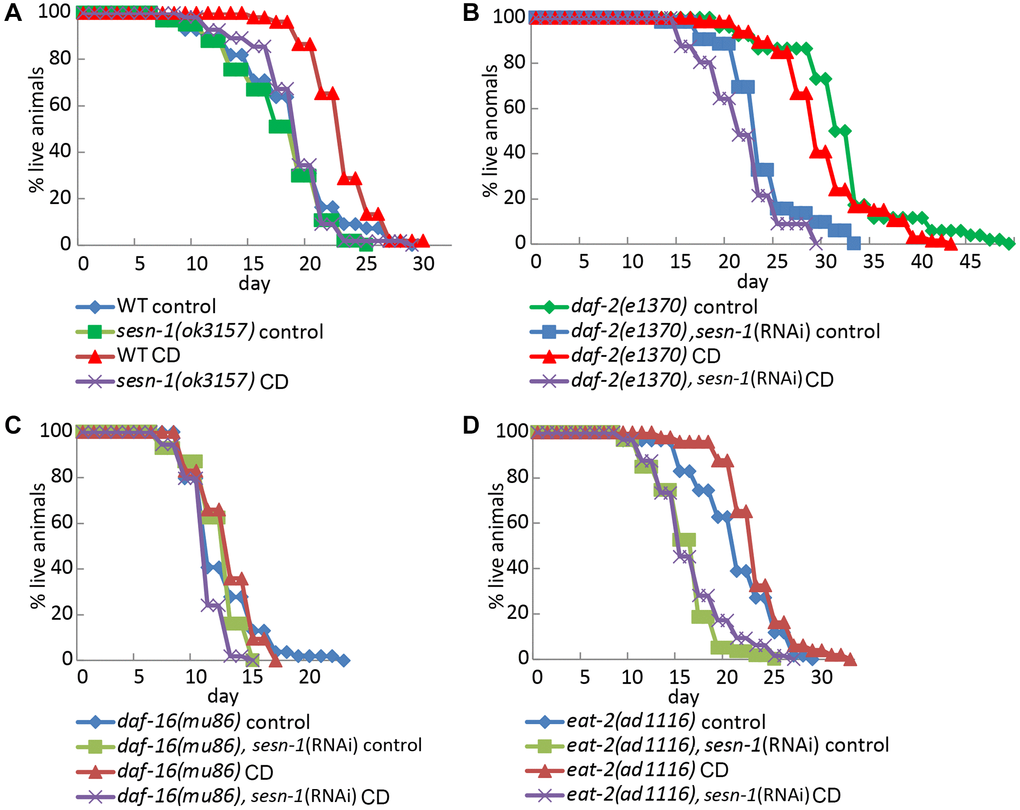
Figure 6. Lifespan regulation by sesn-1 through the daf-16 pathway and in eat-2 mutant under starvation. Analysis of the lifespan of different C. elegans strains: (A) WT and sesn-1(ok3157), (B) daf-2(e1370), (C) daf-16(mu86) and (D) eat-2(ad1116), with and without sesn-1(RNAi), incubated under control or caloric deprivation (CD) conditions.
Table 2. Lifespan extension means analysis under CD for daf-2 (e1370), eat-2 (ad1116), daf-16 (mu86) nematodes with sesn-1 RNAi expression and WT.
| Strain | Sesn-1 gene status | Ortholog | Control mean lifespan ± SEM, days | n | Starvation mean lifespan ± SEM, days | n | Effect vs. control, % | p-value vs. control |
| WT | WT | 18,2 ± 0,64 | 55 | 22,8 ± 0,39 | 52 | +25,3 | <0,0001 | |
| sesn-1(ok3157) | deletion | 17,3 ± 0,56 | 51 | 18,6 ± 0,48 | 55 | +8,7 | 0,4054 | |
| daf-2 (e1370) | WT | IGF1r | 31,8 ± 0,84 | 53 | 29,9 ± 0,62 | 66 | −6,1 | 0,0252 |
| daf-2 (e1370) | RNAi | IGF1r | 23,4 ± 0,56 | 52 | 21,4 ± 0,52 | 56 | −8,7 | 0,0219 |
| eat-2 (ad1116) | WT | 20,8 ± 0,58 | 59 | 23,1 ± 0,5 | 49 | +11,0 | 0,0057 | |
| eat-2 (ad1116) | RNAi | 15,8 ± 0,41 | 59 | 16,3 ± 0,46 | 64 | +3,6 | <0,0001 | |
| daf-16 (mu86) | WT | FOXO family | 12,4 ± 0,41 | 54 | 13,3 ± 0,34 | 53 | +7,5 | >0,9999 |
| daf-16 (mu86) | RNAi | FOXO family | 12,2 ± 0,25 | 69 | 11,0 ± 0,22 | 55 | −9,5 | 0,7069 |
Discussion
Aging is one of humanity’s most pressing challenges, exacerbated by the global rise in life expectancy and the parallel increase in age-related diseases such as cancer, diabetes and neurodegenerative disorders. Understanding the complexities of aging is crucial for mitigating its adverse effects on human well-being and ensuring healthy aging. Notably, the key signaling pathways that govern aging are evolutionary conserved. Model organisms like C. elegans serve as indispensable tools for unravelling the molecular mechanisms underlying aging. CR has emerged as a key physiological intervention that extends both lifespan and healthspan across a wide range of eukaryotic species. In nematodes, CR as well as CD mimic the lifespan-extending effects observed in worms with the eat-2(ad1116) mutation, which significantly restricts food intake [43]. Interestingly, overlaying CD and CR on eat-2 mutants does not further enhance this lifespan extension (Figure 6D and Table 2), suggesting that the eat-2 mutation, CD and CR may share common mechanisms of lifespan regulation.
The central role of mTORC1, a critical nutrient sensor that is deactivated by glucose and amino acid scarcity, is evident in the aging regulation across diverse species from yeast to mice [25]. Both CR and inhibition of mTORC1 similarly extend lifespan, implying a common underlying mechanism [24]. Autophagy activation is a critical mechanism of lifespan extension, as evidenced by the abatement of lifespan extension in animals with inhibited autophagy during CR [41]. In C. elegans, this lifespan enhancement coincides with increased stress tolerance, a benefit attributed to activated autophagy, particularly during food scarcity [52, 53]. Adequate activation of autophagy by Sestrins may underlie the phenomenon of hormesis, the adaptive response to severe stress following exposure to low-level stress, which could also promote lifespan and healthspan extension [53].
Various sensors activated by different nutrients and stress factors likely modulate autophagy, stress response and longevity. Essential proteins, including Sestrins, may integrate these signals and direct them to mTORC1-regulated autophagy, enabling an adaptive response to changing environments. In mammals, glucose shortage and amino acid deprivation increase SESN2 expression via a mechanism mediated by ATF4 [4, 54]. Meanwhile, oxidative stress, DNA damage and hypoxia stimulate Sestrins’ expression through p53, NRF2 and some other transcription factors [37, 55, 56]. Interestingly, in our studies, sesn-1 mRNA levels peaked at 10 hours post-CD initiation but declined by 16 hours, suggesting potential post-transcriptional regulation (data not shown). This could be explained by a decoupling of SESN-1 protein and mRNA expression, with the protein potentially remaining stable despite the reduction in mRNA levels – similar to the dynamics observed for the mammalian SESN2 protein after 24 hours of H2O2 exposure [9]. Such protein stabilization, coupled with its mRNA downregulation during continuous stress, may serve an adaptive function, conserving energy and optimizing recovery when conditions improve. Deletion of sesn-1 significantly attenuated CD-induced lifespan extension, highlighting its critical role in this process (Figure 1) [35, 57]. The slight extension observed in sesn-1-null animals subjected to CD might arise from Sestrin-independent nutrient sensing effects of other unidentified pathways.
Given autophagy’s strong links to longevity and stress mitigation [44], we investigated sesn-1’s involvement in regulating autophagy and its key effector, mTORC1 [41, 51], in the context of CD. Our findings demonstrate that sesn-1 mediates mTORC1 suppression and autophagy activation during food scarcity, potentially driving lifespan extension and stress resilience (Figures 1 and 2A–2C). Failure of sesn-1 (RNAi) worms to activate autophagy properly in response to starvation (Figure 3B) and activate autophagosome formation at the appropriate level in response to chloroquine exposure (Supplementary Figure 2) indicates a remarkable role of sesn-1 in this process. RNA-interference experiments (Figure 6B and Table 2) further support sesn-1’s upstream role in the TORC1 pathway. Our studies (Figure 6C and Table 2) also suggest a possible interconnection between sesn-1 and the daf-16(FOXO) transcription factor, independent of the insulin/IGF pathway. However, these studies should be interpreted with caution and require further evaluation.
Many of Sestrin’s functions appear to be mediated through its interactions with GATOR2. Our study showed that GATOR2-deficient worms failed to exhibit sesn-1-driven lifespan extension during CD, indicating GATOR2’s essential role in this pathway (Figure 4A, 4B and Table 1). Thus, Sestrins may primarily exert their lifespan-regulating and stress-protective effects through modulation of mTORC1 activity and autophagy. This proposition aligns with the studies in D. melanogaster, underscoring the interplay between BCAAs deprivation, dSesn, mTORC1 and autophagy [37]. BCAAs restriction has also been noted to improve metabolic health in humans [58].
Future endeavours targeting sesn-1 deactivation in diverse cell types could shed light on its nuanced roles in regulating longevity and stress response. Elucidating the beneficial role of sesn-1 in nematode lifespan extension has important implications for developing strategies to enhance lifespan and healthspan in humans, given the evolutionary conservation of Sestrins across metazoan species. Targeting Sestrins could pave the way for therapeutic interventions that mimic CR benefits, offering promising strategies for mitigating age-associated diseases and delaying aging.
Materials and Methods
Strains of C. elegans and their maintenance conditions
Strains sourced from the Caenorhabditis Genetics Center (CGC) included: C. elegans Bristol N2 WT strain, RB2325 sesn-1(ok3157)I with deletion of 535bp in exon 3 of the sesn-1 gene, DA2126 (adls2122 (lgg-1p::GFP::LGG-1 + rol-6(su1006)), PD4251(ccIs4251)I, e1282 IV (myo-3p::GFP::LacZ::NLS + myo-3p::mitochondrial GFP + dpy-20(+)), CB1370 daf-2(e1370)III, DA1116 eat-2(ad1116)II, CF1038 daf-16(mu86)I, E. coli OP50 and HT115(DE3) strains. Additionally, IE24589 sesn-1(ie24589), with an MOS-1 transposon insertion in exon 3 of the sesn-1 gene, was kindly provided by Yohann Duverger, Universite Lyon. All strains underwent 8 outcrossing against the Bristol N2 WT strain to mitigate off-target mutations as outlined previously [59]. The C. elegans strains were maintained, synchronous culture was obtained and experiments were performed under standard conditions at 20°C [60].
RNAi-expressing plasmid construction
RNA interference in C. elegans is a method to silence genes by introducing double-stranded RNA, which triggers degradation of a specific endogenous mRNA, effectively inactivating a target gene [61]. Total nematode RNA was extracted using Reagent ExtractRNA (Eurogen, Moscow, Russia) per manufacturer guidelines. Using reverse transcriptase Mint (Eurogen, Moscow, Russia), cDNA was synthesized and the subsequent cDNA fragments were PCR-amplified. The PCR primer pairs are shown in Table 3. The L4440 vector (Addgene, Watertown, MA, USA) was ligated with the respective cDNA fragments at XbaI and BamHI sites to create the RNAi-expressing constructs. Empty vector was used as control.
Table 3. Primer sequences for C. elegans candidate genes.
| Gene | Type | Sequence | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| sesn-1 | F | 5′-agagagtctagaaccatgcacactac-3′ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| R | 5′-agagaggatcctcaatccaaagcctt-3′ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| npp-18 | F | 5′-agatctagagccagcgatatgacaatggcg-3′ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| R | 5′-attggatcctcgggcatggtagatcgaagac-3′ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Y32H12A.8 | F | 5′-agatctagacgatctcatcgaaggtccatcg-3′ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| R | 5′-ataggatccccaccacctgtggcaataagc-3′ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: F: Forward Prime; R: Reverse Primer. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Bacteria-fed RNAi
Briefly, the transformed HT115 strain was grown in LB medium containing tetracycline (12.5 μg/ml) and ampicillin (100 μg/ml) at 37°C with shaking [62]. The bacteria were then plated on ampicillin-containing agarose plates supplemented with 1 mM IPTG and incubated at room temperature for two days. L1 C. elegans were added the following day. For double RNAi experiments, plates were prepared in a similar manner except that the 1:1 mixture of both RNAi bacterial clones was seeded into the plates.
qPCR analysis of mRNA
Synchronized L1 nematodes were placed on bacteria-laden NGM plates. L3 worms were rinsed off the plates with M9, then cleansed three times with PBS. After washing the worms with PBS, they were suspended in the 0.1% Tween-20–PBS solution for 20 minutes to eliminate gut bacteria. The worms were snap-frozen in liquid nitrogen and RNA extraction was conducted using Reagent ExtractRNA (Eurogen, Moscow, Russia). A NanoDrop (Thermo Fisher Scientific Inc, Waltham, MA, USA) was used to determine RNA concentrations and the reverse transcription Mint Kit (Eurogen, Moscow, Russia) was utilized to synthesize cDNA from 3 μg of total RNA. mRNA levels were analyzed using the CFX96 (Bio-Rad Laboratories, Hercules, CA, USA) with qPCRmix-HS SYBR (Eurogen, Moscow, Russia) and assessed using the Bio-Rad CFX Manager software (Bio-Rad Laboratories, Hercules, CA, USA) (Supplementary Figure 3). Topoisomerase I (top-1) mRNA was used as a reference mRNA. The qPCR primer pairs listed in Table 4.
Table 4. Primer sequences for RNAi qPCR validation.
| Gene | Type | Sequence | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| sesn-1 | F | 5′-tccgtgaagcaatttggaac-3′ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| R | 5′-tcgctaccatcattaccacg-3′ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| npp-18 | F | 5′-ttggcgcgttatttgggctc-3′ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| R | 5′-gttcttcggatccattgggattct-3′ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Y32H12A.8 | F | 5′-ccacgacgtcgtcaaacgg-3′ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| R | 5′-gttgcatgccaatcgaggc-3′ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| top-1 | F | 5′-ggcccagaagaagtacgacagactg-3’ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| R | 5′-tcgatggcccaacggaatttc-3′ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: F: Forward Prime; R: Reverse Primer. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Lifespan analysis during CD
Synchronized L1 nematodes were plated on either RNAi plates or empty vector control plates. Upon reaching L4, worms were transferred to RNAi plates, empty vector control plates or plates lacking bactopeptone and bacteria. Viability was assessed daily according to the established protocols [39, 40]. Worms that disappeared or dried on the plate wall were excluded from the analysis.
M9 media, paraquat and hydrogen peroxide survival analysis
To analyze viability in M9 media, L1 nematodes were introduced to 6 cm Petri plates with a thin M9 layer and kept in a 20°C incubator with gentle shaking. Daily aliquots were plated on NGM to count live nematodes. For paraquat and hydrogen peroxide treatments, L4 nematodes were added to 24-well plates containing thin layers of M9 media enriched with either 300 μM paraquat or 4 mM hydrogen peroxide. Viability was determined as previously described.
Fluorescent microscopy analysis of autophagy and muscle degeneration
Autophagosomes in seam cells of the adls2122 strain, expressing the LGG-1::GFP fusion protein, were quantified in both wild-type and sesn-1(RNAi) L3 animals using a Leica DMI4000B inverted microscope (Leica Microsystems, Wetzlar, Germany) at 640× magnification, as previously described [63, 64]. The number of myocyte nuclei labelled with GFP in ccIs4251 strain, which expresses a myo-3 promoter driving a nuclear-targeted GFP-LacZ fusion and myo-3 promoter driving mitochondrially targeted GFP, was analyzed in the same manner for worms at the L4 stage and at the 5-day-old adult stage.
Immunoblotting
Normalized nematode lysates containing 10 mg of protein per sample were subjected to 15% PAGE, as previously described [63–65]. The primary antibodies used for these studies were: anti-GFP (#11814460001, Roche Pharma, Penzberg, Germany), anti-phospho-ribosomal protein S6 (#sc-54279, Santa Cruz Biotechnology, Dallas, TX, USA) and anti-actin (#sc-47778, Santa Cruz Biotechnology, Dallas, TX, USA). The specificity of the anti-phospho-ribosomal protein S6 antibody was confirmed using a blocking peptide (#sc-54279 P, Santa Cruz Biotechnology, Dallas, TX, USA), adhering to the provided protocol. Blot images were acquired using the ChemiDoc Imaging System (Bio-Rad Laboratories, Hercules, CA, USA).
Statistical analysis
Statistical analyses were performed using GraphPad Prism 10 software (GraphPad Software, Boston, MA, USA). One-way ANOVA with Bonferroni correction (p < 0.05) for multiple comparisons was used to assess the statistical significance of differences in mean lifespan across groups in all survival experiments. Lifespan tables show the p-value used to compare the studied groups with their controls. For autophagy and muscle degeneration analyses, Student’s unpaired two-tailed t-test was performed to compare the experimental groups with their respective controls. ns, *, ** and *** correspond to p-values > 0.05, < 0.05, < 0.01 and < 0.001, respectively.
Supplementary Materials
Author Contributions
AOZ performed all studies, PMC and AVB designed the experiments and wrote the paper.
Acknowledgments
We thank Yohann Duverger, Universite Lyon, for the generous gift of IE24589 Sesn-1 deficient strain and Nadusha Pryadilova and Alex Haidurov for help with the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
The work was supported by Grant 17-14-01420 and 23-14-00370 from the Russian Science Foundation, the Welcome Trust ISSF grant and Trinity St. James’s Cancer Institute Cancer Research Stimulus Award (TSJCI Crest Award) to AVB, grant 075-15-2019-1660 from the Russian Federal Research Program for Genetic Technologies Development to PMC.
References
- 1. Velasco-Miguel S, Buckbinder L, Jean P, Gelbert L, Talbott R, Laidlaw J, Seizinger B, Kley N. PA26, a novel target of the p53 tumor suppressor and member of the GADD family of DNA damage and growth arrest inducible genes. Oncogene. 1999; 18:127–37. https://doi.org/10.1038/sj.onc.1202274 [PubMed]
- 2. Budanov AV, Shoshani T, Faerman A, Zelin E, Kamer I, Kalinski H, Gorodin S, Fishman A, Chajut A, Einat P, Skaliter R, Gudkov AV, Chumakov PM, Feinstein E. Identification of a novel stress-responsive gene Hi95 involved in regulation of cell viability. Oncogene. 2002; 21:6017–31. https://doi.org/10.1038/sj.onc.1205877 [PubMed]
- 3. Peeters H, Debeer P, Bairoch A, Wilquet V, Huysmans C, Parthoens E, Fryns JP, Gewillig M, Nakamura Y, Niikawa N, Van de Ven W, Devriendt K. PA26 is a candidate gene for heterotaxia in humans: identification of a novel PA26-related gene family in human and mouse. Hum Genet. 2003; 112:573–80. https://doi.org/10.1007/s00439-003-0917-5 [PubMed]
- 4. Ding B, Parmigiani A, Divakaruni AS, Archer K, Murphy AN, Budanov AV. Sestrin2 is induced by glucose starvation via the unfolded protein response and protects cells from non-canonical necroptotic cell death. Sci Rep. 2016; 6:22538. https://doi.org/10.1038/srep22538 [PubMed]
- 5. Dalina AA, Kovaleva IE, Budanov AV. [Sestrins are Gatekeepers in the Way from Stress to Aging and Disease]. Mol Biol (Mosk). 2018; 52:948–62. https://doi.org/10.1134/S0026898418060046 [PubMed]
- 6. Parmigiani A, Budanov AV. Sensing the Environment Through Sestrins: Implications for Cellular Metabolism. Int Rev Cell Mol Biol. 2016; 327:1–42. https://doi.org/10.1016/bs.ircmb.2016.05.003 [PubMed]
- 7. Haidurov A, Budanov AV. Locked in Structure: Sestrin and GATOR-A Billion-Year Marriage. Cells. 2024; 13:1587. https://doi.org/10.3390/cells13181587 [PubMed]
- 8. Lee JH, Budanov AV, Park EJ, Birse R, Kim TE, Perkins GA, Ocorr K, Ellisman MH, Bodmer R, Bier E, Karin M. Sestrin as a feedback inhibitor of TOR that prevents age-related pathologies. Science. 2010; 327:1223–8. https://doi.org/10.1126/science.1182228 [PubMed]
- 9. Budanov AV, Sablina AA, Feinstein E, Koonin EV, Chumakov PM. Regeneration of peroxiredoxins by p53-regulated sestrins, homologs of bacterial AhpD. Science. 2004; 304:596–600. https://doi.org/10.1126/science.1095569 [PubMed]
- 10. Ding B, Parmigiani A, Yang C, Budanov AV. Sestrin2 facilitates death receptor-induced apoptosis in lung adenocarcinoma cells through regulation of XIAP degradation. Cell Cycle. 2015; 14:3231–41. https://doi.org/10.1080/15384101.2015.1084447 [PubMed]
- 11. Ding B, Haidurov A, Chawla A, Parmigiani A, van de Kamp G, Dalina A, Yuan F, Lee JH, Chumakov PM, Grossman SR, Budanov AV. p53-inducible SESTRINs might play opposite roles in the regulation of early and late stages of lung carcinogenesis. Oncotarget. 2019; 10:6997–7009. https://doi.org/10.18632/oncotarget.27367 [PubMed]
- 12. Kozak J, Jonak K. Association between the antioxidant properties of SESN proteins and anti-cancer therapies. Amino Acids. 2023; 55:835–51. https://doi.org/10.1007/s00726-023-03281-6 [PubMed]
- 13. Haidurov A, Zheltukhin AO, Snezhkina AV, Krasnov GS, Kudryavtseva AV, Budanov AV. p53-regulated SESN1 and SESN2 regulate cell proliferation and cell death through control of STAT3. Cell Commun Signal. 2025; 23:105. https://doi.org/10.1186/s12964-025-02104-3 [PubMed]
- 14. Budanov AV, Karin M. p53 target genes sestrin1 and sestrin2 connect genotoxic stress and mTOR signaling. Cell. 2008; 134:451–60. https://doi.org/10.1016/j.cell.2008.06.028 [PubMed]
- 15. Lee JH, Budanov AV, Talukdar S, Park EJ, Park HL, Park HW, Bandyopadhyay G, Li N, Aghajan M, Jang I, Wolfe AM, Perkins GA, Ellisman MH, et al. Maintenance of metabolic homeostasis by Sestrin2 and Sestrin3. Cell Metab. 2012; 16:311–21. https://doi.org/10.1016/j.cmet.2012.08.004 [PubMed]
- 16. Sablina AA, Budanov AV, Ilyinskaya GV, Agapova LS, Kravchenko JE, Chumakov PM. The antioxidant function of the p53 tumor suppressor. Nat Med. 2005; 11:1306–13. https://doi.org/10.1038/nm1320 [PubMed]
- 17. Liu GY, Sabatini DM. mTOR at the nexus of nutrition, growth, ageing and disease. Nat Rev Mol Cell Biol. 2020; 21:183–203. https://doi.org/10.1038/s41580-019-0199-y [PubMed]
- 18. Bialik S, Dasari SK, Kimchi A. Autophagy-dependent cell death - where, how and why a cell eats itself to death. J Cell Sci. 2018; 131:jcs215152. https://doi.org/10.1242/jcs.215152 [PubMed]
- 19. Powers RW
3rd , Kaeberlein M, Caldwell SD, Kennedy BK, Fields S. Extension of chronological life span in yeast by decreased TOR pathway signaling. Genes Dev. 2006; 20:174–84. https://doi.org/10.1101/gad.1381406 [PubMed] - 20. Vellai T, Takacs-Vellai K, Zhang Y, Kovacs AL, Orosz L, Müller F. Genetics: influence of TOR kinase on lifespan in C. elegans. Nature. 2003; 426:620. https://doi.org/10.1038/426620a [PubMed]
- 21. Kapahi P, Zid BM, Harper T, Koslover D, Sapin V, Benzer S. Regulation of lifespan in Drosophila by modulation of genes in the TOR signaling pathway. Curr Biol. 2004; 14:885–90. https://doi.org/10.1016/j.cub.2004.03.059 [PubMed]
- 22. Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, Miller RA. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009; 460:392–5. https://doi.org/10.1038/nature08221 [PubMed]
- 23. Lyu YX, Fu Q, Wilczok D, Ying K, King A, Antebi A, Vojta A, Stolzing A, Moskalev A, Georgievskaya A, Maier AB, Olsen A, Groth A, et al. Longevity biotechnology: bridging AI, biomarkers, geroscience and clinical applications for healthy longevity. Aging (Albany NY). 2024; 16:12955–76. https://doi.org/10.18632/aging.206135 [PubMed]
- 24. Hansen M, Taubert S, Crawford D, Libina N, Lee SJ, Kenyon C. Lifespan extension by conditions that inhibit translation in Caenorhabditis elegans. Aging Cell. 2007; 6:95–110. https://doi.org/10.1111/j.1474-9726.2006.00267.x [PubMed]
- 25. Johnson SC, Rabinovitch PS, Kaeberlein M. mTOR is a key modulator of ageing and age-related disease. Nature. 2013; 493:338–45. https://doi.org/10.1038/nature11861 [PubMed]
- 26. Pan H, Finkel T. Key proteins and pathways that regulate lifespan. J Biol Chem. 2017; 292:6452–60. https://doi.org/10.1074/jbc.R116.771915 [PubMed]
- 27. Murphy CT, Hu PJ. Insulin/insulin-like growth factor signaling in C. elegans. WormBook. 2013; 1–43. https://doi.org/10.1895/wormbook.1.164.1 [PubMed]
- 28. Gross DN, van den Heuvel AP, Birnbaum MJ. The role of FoxO in the regulation of metabolism. Oncogene. 2008; 27:2320–36. https://doi.org/10.1038/onc.2008.25 [PubMed]
- 29. Zaarur N, Desevin K, Mackenzie J, Lord A, Grishok A, Kandror KV. ATGL-1 mediates the effect of dietary restriction and the insulin/IGF-1 signaling pathway on longevity in C. elegans. Mol Metab. 2019; 27:75–82. https://doi.org/10.1016/j.molmet.2019.07.001 [PubMed]
- 30. Huayta J, Crapster JP, San-Miguel A. Endogenous DAF-16 spatiotemporal activity quantitatively predicts lifespan extension induced by dietary restriction. Commun Biol. 2023; 6:203. https://doi.org/10.1038/s42003-023-04562-2 [PubMed]
- 31. Saxton RA, Sabatini DM. mTOR Signaling in Growth, Metabolism, and Disease. Cell. 2017; 168:960–76. https://doi.org/10.1016/j.cell.2017.02.004 [PubMed]
- 32. Parmigiani A, Nourbakhsh A, Ding B, Wang W, Kim YC, Akopiants K, Guan KL, Karin M, Budanov AV. Sestrins inhibit mTORC1 kinase activation through the GATOR complex. Cell Rep. 2014; 9:1281–91. https://doi.org/10.1016/j.celrep.2014.10.019 [PubMed]
- 33. Chantranupong L, Wolfson RL, Orozco JM, Saxton RA, Scaria SM, Bar-Peled L, Spooner E, Isasa M, Gygi SP, Sabatini DM. The Sestrins interact with GATOR2 to negatively regulate the amino-acid-sensing pathway upstream of mTORC1. Cell Rep. 2014; 9:1–8. https://doi.org/10.1016/j.celrep.2014.09.014 [PubMed]
- 34. Yang YL, Loh KS, Liou BY, Chu IH, Kuo CJ, Chen HD, Chen CS. SESN-1 is a positive regulator of lifespan in Caenorhabditis elegans. Exp Gerontol. 2013; 48:371–9. https://doi.org/10.1016/j.exger.2012.12.011 [PubMed]
- 35. Zheltukhin AO, Ilyinskaya GV, Budanov AV, Chumakov PM. Some Phenotypic Characteristics of Nematode Caenorhabditis Elegans Strains with Defective Functions of the Sestrin (cSesn) Gene. Biomed Pharmacol J. 2018; 11:759–67. https://doi.org/10.13005/bpj/1430
- 36. Juricic P, Grönke S, Partridge L. Branched-Chain Amino Acids Have Equivalent Effects to Other Essential Amino Acids on Lifespan and Aging-Related Traits in Drosophila. J Gerontol A Biol Sci Med Sci. 2020; 75:24–31. https://doi.org/10.1093/gerona/glz080 [PubMed]
- 37. Lu J, Temp U, Müller-Hartmann A, Esser J, Grönke S, Partridge L. Sestrin is a key regulator of stem cell function and lifespan in response to dietary amino acids. Nat Aging. 2021; 1:60–72. https://doi.org/10.1038/s43587-020-00001-7 [PubMed]
- 38. Haidurov A, Budanov AV. Sestrin family - the stem controlling healthy ageing. Mech Ageing Dev. 2020; 192:111379. https://doi.org/10.1016/j.mad.2020.111379 [PubMed]
- 39. Lee GD, Wilson MA, Zhu M, Wolkow CA, de Cabo R, Ingram DK, Zou S. Dietary deprivation extends lifespan in Caenorhabditis elegans. Aging Cell. 2006; 5:515–24. https://doi.org/10.1111/j.1474-9726.2006.00241.x [PubMed]
- 40. Kaeberlein TL, Smith ED, Tsuchiya M, Welton KL, Thomas JH, Fields S, Kennedy BK, Kaeberlein M. Lifespan extension in Caenorhabditis elegans by complete removal of food. Aging Cell. 2006; 5:487–94. https://doi.org/10.1111/j.1474-9726.2006.00238.x [PubMed]
- 41. Hansen M, Chandra A, Mitic LL, Onken B, Driscoll M, Kenyon C. A role for autophagy in the extension of lifespan by dietary restriction in C. elegans. PLoS Genet. 2008; 4:e24. https://doi.org/10.1371/journal.pgen.0040024 [PubMed]
- 42. Jia K, Chen D, Riddle DL. The TOR pathway interacts with the insulin signaling pathway to regulate C. elegans larval development, metabolism and life span. Development. 2004; 131:3897–906. https://doi.org/10.1242/dev.01255 [PubMed]
- 43. Lakowski B, Hekimi S. The genetics of caloric restriction in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 1998; 95:13091–6. https://doi.org/10.1073/pnas.95.22.13091 [PubMed]
- 44. Mizushima N. Autophagy: process and function. Genes Dev. 2007; 21:2861–73. https://doi.org/10.1101/gad.1599207 [PubMed]
- 45. Li Y, Tian X, Luo J, Bao T, Wang S, Wu X. Molecular mechanisms of aging and anti-aging strategies. Cell Commun Signal. 2024; 22:285. https://doi.org/10.1186/s12964-024-01663-1 [PubMed]
- 46. Palmisano NJ, Meléndez A. Detection of Autophagy in Caenorhabditis elegans Using GFP::LGG-1 as an Autophagy Marker. Cold Spring Harb Protoc. 2016; 2016:pdb.prot086496. https://doi.org/10.1101/pdb.prot086496 [PubMed]
- 47. Klionsky DJ, Abdelmohsen K, Abe A, Abedin MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD, Adeli K, Adhihetty PJ, Adler SG, Agam G, et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy. 2016; 12:1–222. https://doi.org/10.1080/15548627.2015.1100356 [PubMed]
- 48. Baxi K, Ghavidel A, Waddell B, Harkness TA, de Carvalho CE. Regulation of Lysosomal Function by the DAF-16 Forkhead Transcription Factor Couples Reproduction to Aging in Caenorhabditis elegans. Genetics. 2017; 207:83–101. https://doi.org/10.1534/genetics.117.204222 [PubMed]
- 49. Segalés J, Perdiguero E, Serrano AL, Sousa-Victor P, Ortet L, Jardí M, Budanov AV, Garcia-Prat L, Sandri M, Thomson DM, Karin M, Hee Lee J, Muñoz-Cánoves P. Sestrin prevents atrophy of disused and aging muscles by integrating anabolic and catabolic signals. Nat Commun. 2020; 11:189. https://doi.org/10.1038/s41467-019-13832-9 [PubMed]
- 50. Budanov AV. SESTRINs regulate mTORC1 via RRAGs: The riddle of GATOR. Mol Cell Oncol. 2015; 2:e997113. https://doi.org/10.1080/23723556.2014.997113 [PubMed]
- 51. Zhou B, Kreuzer J, Kumsta C, Wu L, Kamer KJ, Cedillo L, Zhang Y, Li S, Kacergis MC, Webster CM, Fejes-Toth G, Naray-Fejes-Toth A, Das S, et al. Mitochondrial Permeability Uncouples Elevated Autophagy and Lifespan Extension. Cell. 2019; 177:299–314.e16. https://doi.org/10.1016/j.cell.2019.02.013 [PubMed]
- 52. Kang C, You YJ, Avery L. Dual roles of autophagy in the survival of Caenorhabditis elegans during starvation. Genes Dev. 2007; 21:2161–71. https://doi.org/10.1101/gad.1573107 [PubMed]
- 53. Kumsta C, Hansen M. Hormetic heat shock and HSF-1 overexpression improve C. elegans survival and proteostasis by inducing autophagy. Autophagy. 2017; 13:1076–7. https://doi.org/10.1080/15548627.2017.1299313 [PubMed]
- 54. Ye J, Palm W, Peng M, King B, Lindsten T, Li MO, Koumenis C, Thompson CB. GCN2 sustains mTORC1 suppression upon amino acid deprivation by inducing Sestrin2. Genes Dev. 2015; 29:2331–6. https://doi.org/10.1101/gad.269324.115 [PubMed]
- 55. Wolfson RL, Chantranupong L, Saxton RA, Shen K, Scaria SM, Cantor JR, Sabatini DM. Sestrin2 is a leucine sensor for the mTORC1 pathway. Science. 2016; 351:43–8. https://doi.org/10.1126/science.aab2674 [PubMed]
- 56. Kimball SR, Gordon BS, Moyer JE, Dennis MD, Jefferson LS. Leucine induced dephosphorylation of Sestrin2 promotes mTORC1 activation. Cell Signal. 2016; 28:896–906. https://doi.org/10.1016/j.cellsig.2016.03.008 [PubMed]
- 57. Kovaleva IE, Tokarchuk AV, Zheltukhin AO, Dalina AA, Safronov GG, Evstafieva AG, Lyamzaev KG, Chumakov PM, Budanov AV. Mitochondrial localization of SESN2. PLoS One. 2020; 15:e0226862. https://doi.org/10.1371/journal.pone.0226862 [PubMed]
- 58. Fontana L, Cummings NE, Arriola Apelo SI, Neuman JC, Kasza I, Schmidt BA, Cava E, Spelta F, Tosti V, Syed FA, Baar EL, Veronese N, Cottrell SE, et al. Decreased Consumption of Branched-Chain Amino Acids Improves Metabolic Health. Cell Rep. 2016; 16:520–30. https://doi.org/10.1016/j.celrep.2016.05.092 [PubMed]
- 59. Wang D, Cao M, Dinh J, Dong Y. Methods for creating mutations in C. elegans that extend lifespan. Methods Mol Biol. 2013; 1048:65–75. https://doi.org/10.1007/978-1-62703-556-9_6 [PubMed]
- 60. Stiernagle T. Maintenance of C. elegans. WormBook. 2006; 1–11. https://doi.org/10.1895/wormbook.1.101.1 [PubMed]
- 61. Conte D
Jr , MacNeil LT, Walhout AJM, Mello CC. RNA Interference in Caenorhabditis elegans. Curr Protoc Mol Biol. 2015; 109:26.3.1–26.3.30. https://doi.org/10.1002/0471142727.mb2603s109 [PubMed] - 62. Timmons L, Court DL, Fire A. Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans. Gene. 2001; 263:103–12. https://doi.org/10.1016/s0378-1119(00)00579-5 [PubMed]
- 63. Palmisano NJ, Meléndez A. Detection of Autophagy in Caenorhabditis elegans by Western Blotting Analysis of LGG-1. Cold Spring Harb Protoc. 2016; 2016:pdb.prot086512. https://doi.org/10.1101/pdb.prot086512 [PubMed]
- 64. Meléndez A, Tallóczy Z, Seaman M, Eskelinen EL, Hall DH, Levine B. Autophagy genes are essential for dauer development and life-span extension in C. elegans. Science. 2003; 301:1387–91. https://doi.org/10.1126/science.1087782 [PubMed]
- 65. Djeddi A, Michelet X, Culetto E, Alberti A, Barois N, Legouis R. Induction of autophagy in ESCRT mutants is an adaptive response for cell survival in C. elegans. J Cell Sci. 2012; 125:685–94. https://doi.org/10.1242/jcs.091702 [PubMed]