



Introduction
Aging is a complex process characterized by progressive physical, functional, and physiological declines [1]. The liver, a vital organ responsible for maintaining homeostasis through various metabolic pathways, undergoes significant age-related morphological and functional changes with age [2]. These alterations can lead to the deterioration of liver function, increasing the risk of various diseases and negatively affecting overall health [3–6]. Therefore, efficient strategies to mitigate the effects of aging on the liver are crucial for promoting health and longevity.
Calorie restriction (CR), a well-established anti-aging intervention, has shown promise in modulating the aging process by regulating inflammation and metabolism [7, 8]. Several studies have reported beneficial effects of CR on liver metabolism and inflammation, with significant improvements in gene expression profiles and lipid accumulation associated with hepatic aging [9–15]. CR has been consistently recognized as the golden standard for anti-aging interventions, demonstrating robust effects on lifespan and health span across species. However, the precise mechanisms underlying the effects of CR on age-related changes in the liver remain unclear.
Recent research has highlighted the intricate interplay between metabolism and circadian rhythms. Circadian regulation of enzymes involved in metabolite synthesis, such as fatty acids and cholesterol, demonstrates the profound influence of circadian rhythms on metabolic gene expression [16, 17]. Genetic alterations in core clock genes can disrupt metabolic functions, leading to impaired gluconeogenesis, hepatic steatosis, obesity, and abnormal lipid and glucose metabolism [18–20]. Age-related declines in circadian rhythms have been linked to disruptions in metabolic tissue homeostasis [21–23], particularly in the liver [24–26]. Our previous study reported that aging induces dysregulation of circadian genes such as Nr1d1, suggesting a close relationship between the aging process and circadian clock dysfunction [27].
Lipid metabolism plays a central role in the aging process, with lipid accumulation exerting detrimental effects on cells and organs, contributing to age-related diseases and reduced lifespan [28–30]. Age-related changes in lipid composition and accumulation have been implicated in metabolic dysfunction-associated fatty liver disease (MAFLD), which increases in prevalence with age [31]. Lipid metabolism dysfunction accelerates the aging process [32], with several studies reporting increased hepatic lipid accumulation during aging [15, 33, 34]. Therefore, identifying key regulators linking circadian rhythms and lipid metabolism in aging contexts is crucial for developing effective healthy aging strategies.
Bioinformatics and systems biology approaches offer valuable tools for unraveling aging mechanisms and identifying critical aging biomarkers [35, 36]. These tools integrate statistical analyses, mathematical models, and pathway/network construction to provide comprehensive understanding of aging. Various studies using biological big data have shown metabolic changes during aging [37–41]. However, many bioinformatics studies remain at the systemic level and lack detailed exploration of specific mechanisms through which anti-aging interventions modulate aging. There is a pressing need for studies that bridge bioinformatics and systems biology with targeted biological experiments to elucidate the molecular underpinnings of aging and the effects of anti-aging interventions.
Our previous studies extensively characterized the beneficial effects of CR on various organs during aging [7, 14, 42]. Building on these findings, the study aimed to investigate age-related gene expression changes in the liver and identify key gene candidates modulated by CR. We employed an integrated approach combining RNA-seq analysis, bioinformatics tools, and targeted biological experiments. By focusing on metabolic homeostasis in the liver, we sought novel insights into molecular mechanisms underlying hepatic aging and the potential of CR to mitigate these effects. We hypothesized that CR counteracts age-related dysregulation of key circadian and metabolic regulators, thereby maintaining metabolic homeostasis in the aging liver. Through this investigation, we discovered that circadian and metabolic regulators Nr1d1 and Pparα were modulated by CR during aging.
Results
Transcriptomic analysis of gene expression changes during aging and CR
We developed an integrated approach combining bioinformatics, systems biology, and empirical biological analyses to investigate aging and CR effects on gene expression in the liver (Figure 1). Liver tissues from young rats (Young group), aged rats (Old group), and aged rats treated with CR (Old-CR group) were harvested for RNA-Seq analysis. We compared mRNA expression levels between the young and old groups (Old vs. Young) and between the old group and old-CR groups (Old-CR vs. Old) to identify differentially expressed genes (DEGs) (Supplementary Table 1). The Old vs. Young comparison revealed 599 upregulated genes and 578 downregulated genes (Figure 2A). In the Old-CR vs. Old comparison, 459 genes were upregulated, and 663 genes were downregulated (Figure 2B).
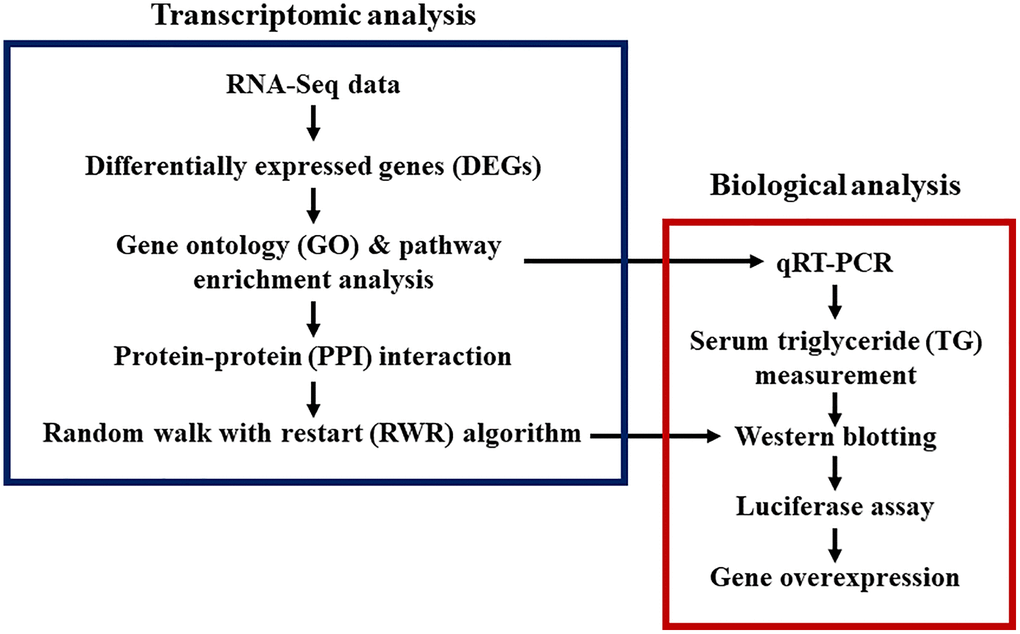
Figure 1. Study designs for transcriptomic and biological analyses of aging and CR. Bioinformatics, systems biology, and biological experiments were combined.
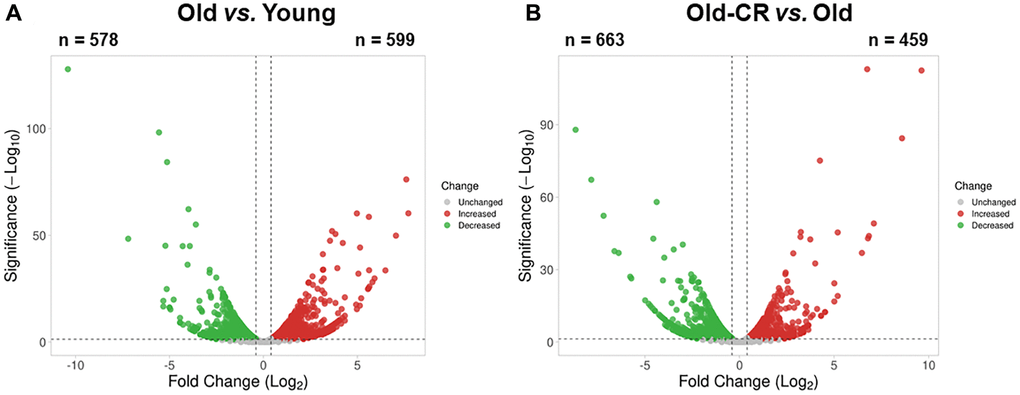
Figure 2. Aging-related differentially expressed genes from RNA-Seq data of the (A) Old vs. Young and (B) Old-CR vs. Old datasets from SD rats. Aging upregulated 599 genes and downregulated 578 genes, while CR in aged mice upregulated 459 genes and downregulated 663 genes. Red dots represent upregulated genes between the two groups, whereas green dots represent downregulated genes. Gray dots represent genes with no changes between the two groups. The DEG criteria were |FC|>1.5 and p < 0.05. Old-CR, old group under CR.
To elucidate DEG functions, we analyzed all genes using the Database for Annotation, Visualization, and Integrated Discovery (DAVID) across five categories: biological processes (BP), cellular components (CC), molecular functions (MF), Kyoto Encyclopedia of Genes and Genomes (KEGG), and Reactome. The top 30 terms based on p-values are listed, to provide insights into the gene functions affected by aging and CR (Supplementary Table 2).
Our results revealed significant changes across various metabolic processes in both Old vs. Young and Old-CR vs. Old datasets. Prevalent BP terms included “aging,” “steroid metabolic process,” “triglyceride metabolic process,” and “xenobiotic metabolic process” (Figure 3A). Key circadian rhythm terms included “circadian rhythm” and “regulation of circadian rhythm,” while immune response terms included “response to lipopolysaccharide”.
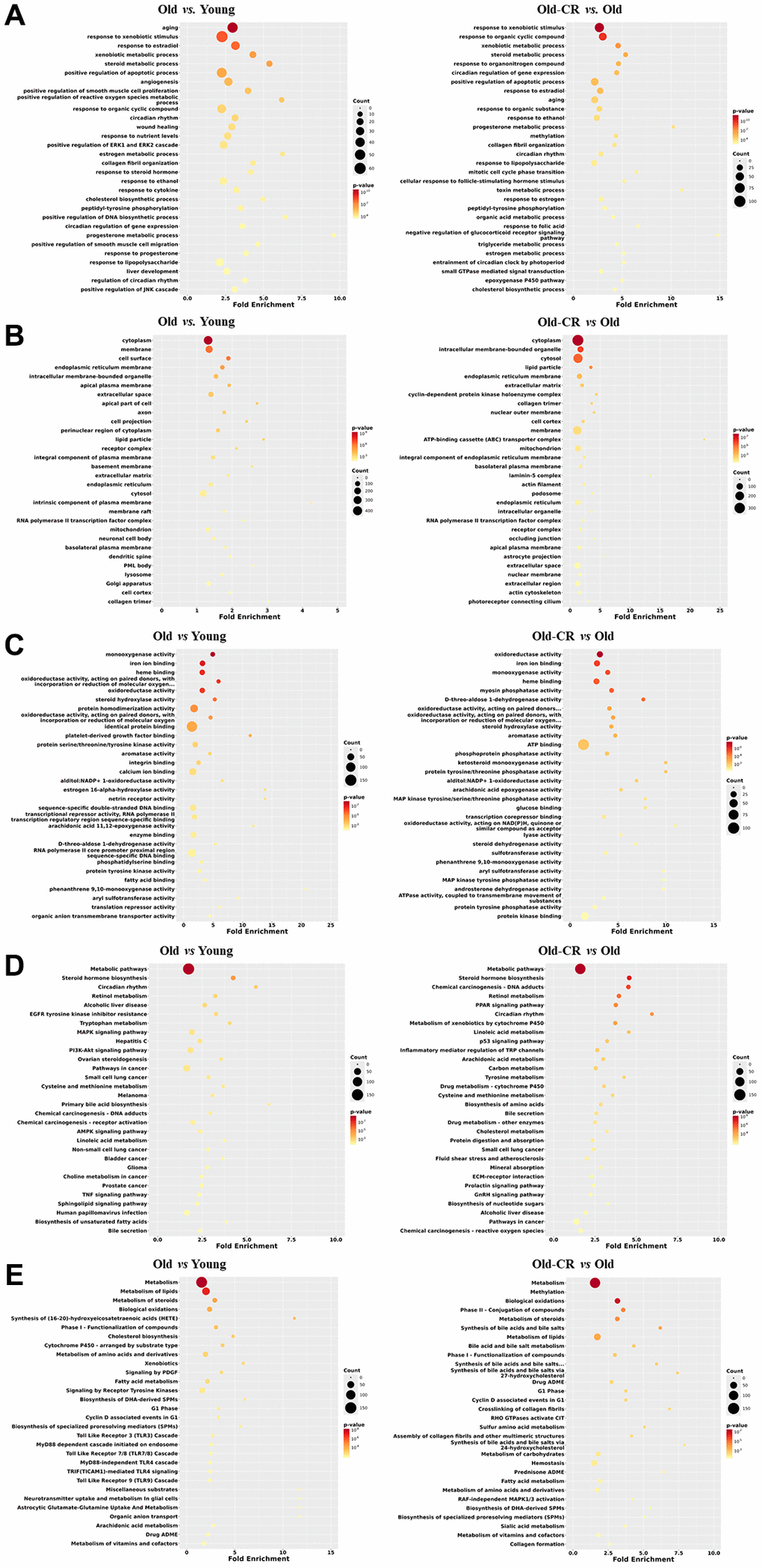
Figure 3. Top 30 enriched GO or pathway terms of the DEGs in the Old vs. Young and Old-CR vs. Old datasets from SD rats. (A) BP, (B) CC, (C) MF, (D) KEGG, and (E) Reactome data. Gene ontology analysis revealed significant alterations in metabolic processes, circadian rhythm, and inflammation (BP); lipid particles, collagen, and mitochondria (CC); and energy/lipid metabolism enzymes (MF). Pathway analysis identified changes in AMPK/PPAR signaling and circadian rhythm (KEGG), with distinct inflammatory signatures in aging and extracellular matrix responses in CR (Reactome). The criterion for a significant term was p < 0.05. Counts indicate the number of genes related to each term in the analysis. The fold enrichment indicates how much a particular GO term or pathway is overrepresented in the list of genes compared to that expected by chance. Old-CR, old group under CR.
CC terms such as “lipid particle,” “collagen trimer,” and “mitochondrion,” were altered in both datasets (Figure 3B). Key MF terms associated with energy metabolism-related “oxidoreductase” and “monooxygenase” and lipid metabolism-related “steroid hydroxylase activity” and “fatty acid binding” (Figure 3C).
KEGG analysis revealed changes in “AMPK signaling pathway,” “PPAR (Peroxisome proliferator-activated receptors) signaling pathway,” and “steroid hormone biosynthesis,” indicating strong associations with lipid metabolism and circadian rhythm-related terms were also identified (Figure 3D). Reactome terms such as “metabolism,” “metabolism of lipids/steroids,” and “fatty acid metabolism,” were prominent in both datasets. The Old vs. Young dataset showed a higher prevalence of terms related to toll-like receptor signaling, whereas the Old-CR vs. Old dataset was enriched in terms related to the extracellular matrix (ECM) terms, such as “crosslinking of collagen fibrils” and “collagen formation” (Figure 3E).
These results suggest that genes associated with inflammation, metabolism, circadian rhythms, and the ECM exhibited differential expression patterns in aging and CR. Consistent changes in metabolism and circadian rhythm-related GOs and pathways highlight their significant interplay in the aging liver, underscoring their critical roles in liver responses to aging and CR interventions.
CR modulation of aging-related changes in lipid metabolism-related gene expression
To explore CR intervention in age-induced changes, we examined DEG expression patterns from both datasets (Supplementary Table 3). Specifically, we conducted GO and pathway analyses focusing on genes upregulated in the Old vs. Young but downregulated in the Old-CR vs. Old (Table 1). BP terms related to fatty acid, lipid, and cholesterol steroid processes and MF terms, including palmitoyl-CoA 9-desaturase and stearoyl-CoA 9-desaturase activities were identified. Metabolic pathways such as the AMPK and PPAR signaling pathways, lipid metabolism, and fatty acid metabolism were altered.
Table 1. GO terms and pathways of genes that were upregulated in the Old vs. Young but downregulated in the Old-CR vs. Old datasets.
| Category | Term | Fold enrichment | p-value | Genes |
| BP | positive regulation of apoptotic process | 4.8095 | 1.60E-05 | MOAP1, TNFRSF12A, SRC, ZBTB16, PDCD5, PTGS2, PPP2CA, CASP12, UBD, ALDH1A1, HMOX1, TSPO, CYP1B1 |
| fatty acid biosynthetic process | 16.37276596 | 2.37E-04 | ACLY, SCD, FASN, SCD2, ACACB | |
| response to organonitrogen compound | 10.68777778 | 0.001206086 | CASP12, UBD, CCNG1, PTGS2, AACS | |
| acetyl-CoA metabolic process | 51.30133333 | 0.001440356 | ACLY, FASN, ACACB | |
| peptidyl-tyrosine phosphorylation | 9.271325301 | 0.002040379 | EFEMP1, SPINK1, SRC, PDGFB, EPHA3 | |
| response to xenobiotic stimulus | 3.129286506 | 0.002650987 | CASP12, SRC, ALDH1A1, PDGFB, HMOX1, TSPO, MVD, CYP4A2, PTGS2, ACACB, AACS | |
| response to fatty acid | 13.38295652 | 0.003241543 | SCD, SRC, SCD2, PTGS2 | |
| regulation of cell-cell adhesion | 28.857 | 0.00466156 | SRC, ADAM8, ZDHHC2 | |
| response to organic cyclic compound | 3.800098765 | 0.005063243 | G6PD, LUM, ALDH1A1, PDGFB, CYP1B1, PTGS2, ACACB, AACS | |
| positive regulation of ERK1 and ERK2 cascade | 4.397257143 | 0.005141411 | CCL21, SRC, C5AR2, TRPV4, PDGFB, GAS6, FGF21 | |
| inflammatory response | 3.439195531 | 0.008574019 | CXCL9, CCL21, ELF3, C5AR2, FASN, ADAM8, CXCL1, PTGS2 | |
| negative regulation of cell proliferation | 3.01772549 | 0.009810343 | ZBTB16, PDCD5, ADORA1, CYP1B1, HMOX1, CD9, CDH13, SLIT3, PTGS2 | |
| lipid biosynthetic process | 19.238 | 0.010367306 | ACLY, SCD, FASN | |
| white fat cell differentiation | 19.238 | 0.010367306 | SCD, PNPLA3, AACS | |
| positive regulation of protein kinase B signaling | 5.785864662 | 0.010820854 | CCL21, SRC, GDF15, ADAM8, GAS6 | |
| positive regulation of DNA biosynthetic process | 17.75815385 | 0.012106014 | SRC, PDGFB, CYP1B1 | |
| neutrophil chemotaxis | 8.208213333 | 0.012599584 | LGALS3, CXCL9, CCL21, CXCL1 | |
| positive regulation of phosphatidylinositol 3-kinase activity | 15.3904 | 0.015934563 | CCL21, SRC, PDGFB | |
| activation of protein kinase B activity | 15.3904 | 0.015934563 | SRC, PDGFB, GAS6 | |
| positive regulation of MAP kinase activity | 7.417060241 | 0.016521383 | SRC, PDGFB, ADAM8, TPD52L1 | |
| response to ethanol | 4.014886957 | 0.016714469 | G6PD, SPINK1, CASP12, GSN, ALDH1A1, AACS | |
| cell chemotaxis | 7.242541176 | 0.017596824 | CCL21, ARHGEF16, PDGFB, CXCL1 | |
| monocyte chemotaxis | 14.4285 | 0.018017922 | LGALS3, CCL21, PDGFB | |
| endothelial cell-cell adhesion | 102.6026667 | 0.019213353 | CYP1B1, THBS4 | |
| positive regulation of smooth muscle cell proliferation | 6.917033708 | 0.019862531 | PDGFB, HMOX1, CDH13, PTGS2 | |
| positive regulation of protein tyrosine kinase activity | 13.57976471 | 0.020209789 | TRPV4, PDGFB, GAS6 | |
| actin filament organization | 4.607904192 | 0.023008755 | GSN, TRPV4, MYO7B, RHOC, TLE6 | |
| positive regulation of reactive oxygen species metabolic process | 12.4787027 | 0.023694306 | PDGFB, CYP1B1, TSPO | |
| aging | 3.131767442 | 0.024104355 | CDKN1C, CASP12, GSN, CNP, TSPO, PTGS2, DCN | |
| antimicrobial humoral immune response mediated by antimicrobial peptide | 6.346556701 | 0.024854734 | LGALS3, CXCL9, CCL21, CXCL1 | |
| positive regulation of glomerular filtration | 76.952 | 0.025536074 | PDGFB, GAS6 | |
| monounsaturated fatty acid biosynthetic process | 76.952 | 0.025536074 | SCD, SCD2 | |
| adrenal gland development | 11.5428 | 0.027405876 | CDKN1C, CYP1B1, TSPO | |
| positive regulation of JNK cascade | 6.095207921 | 0.027581281 | CCL21, TRPV4, TPD52L1, ANKRD6 | |
| immune system process | 11.26126829 | 0.028691809 | IGHM, IRF7 | |
| cholesterol biosynthetic process | 10.99314286 | 0.030001514 | G6PD, MVD, LSS | |
| positive regulation of inflammatory response | 5.863009524 | 0.030460944 | TRPV4, NKG7, ADAM8, IL17RB | |
| fructose catabolic process | 61.5616 | 0.031818362 | ALDH1A1, ALDH1A7 | |
| positive regulation of smooth muscle cell migration | 10.26026667 | 0.03406973 | SRC, PDGFB, CYP1B1 | |
| response to nutrient levels | 4.071534392 | 0.034078992 | SPINK1, SRC, FASN, ACACB, FGF21 | |
| cell adhesion | 2.88055615 | 0.034278073 | TNFRSF12A, SRC, CYP1B1, CD9, CCDC141, MPDZ, THBS4 | |
| positive regulation of I-kappaB kinase/NF-kappaB signaling | 3.966597938 | 0.036959971 | CCL21, UBD, HMOX1, RHOC, TMEM106A | |
| response to iron (III) ion | 51.30133333 | 0.038060473 | G6PD, CASP12 | |
| kidney development | 3.926122449 | 0.038150589 | CDKN1C, ALDH1A1, RHPN1, CYP4A2, DCN | |
| negative regulation of dendritic cell apoptotic process | 43.97257143 | 0.044262663 | CCL21, GAS6 | |
| cellular response to glucose stimulus | 4.88584127 | 0.048049626 | PPP2CA, GAS6, AACS, FGF21 | |
| chemokine-mediated signaling pathway | 8.394763636 | 0.049034302 | CXCL9, CCL21, CXCL1 | |
| CC | extracellular space | 2.208098626 | 5.51E-04 | WFDC21, CXCL9, SPINK1, GSN, CCL21, CNP, GDF15, LUM, PDGFB, MUP4, CTSW, CXCL1, PLA2G7, THBS4, DCN, LGALS3, PROCR, EFEMP1, PODNL1, CDH13, SLIT3, GAS6, FGF21 |
| cytoplasm | 1.393257601 | 0.00602873 | CDKN1C, LGALS3, EFEMP1, CASP12, ME1, ANXA8, CLIC2, SULT4A1, HTATIP2, ANKRD6, CDKL2, G6PD, SPINK1, MYO7B, DUSP26, IL17RB, TLE6, ACLY, EVC, ALDH1A1, IRF7, CDH13, ADAM8, GAS6, CNP, SRC, PDGFB, DCUN1D2, PTGS2, PLA2G7, PPP2CA, UBD, CCDC141, MPDZ, PDLIM4, FGF21, NEK8, MOAP1, GSN, GDF15, MOK, PRSS36, TPD52L1, KY, SBK1, FASN, CCNG1, PNPLA3, B9D1, PNPLA5 | |
| caveola | 6.761153054 | 0.02109186 | SRC, HMOX1, CDH13, PTGS2 | |
| podosome | 13.2405914 | 0.0212081 | GSN, SRC, ADAM8 | |
| membrane | 1.41390075 | 0.033419567 | POPDC2, ADAMDEC1, CNP, SRC, C5AR2, PDGFB, CYP4A2, SLC2A5, ZDHHC2, ADGRG2, SCD2, HMOX1, CYP1B1, SLIT3, TAS1R1, CCDC141, CLIC2, CSMD1, GRIA3, ENTPD7, SYT15, RHOC, IL17RB, PROCR, SCD, TRPV4, PNPLA3, CD9, PNPLA5, B9D1, PLP2, TMEM106A | |
| extracellular region | 1.943573049 | 0.043080377 | LGALS3, WFDC21, GSN, GDF15, PDGFB, CD9, PRSS36, GAS6, ZP2, CHRDL1, DCN, THBS4 | |
| MF | growth factor activity | 7.79222973 | 2.71E-04 | EFEMP1, GDF15, PDGFB, CXCL1, GAS6, THBS4, FGF21 |
| protein homodimerization activity | 2.678861789 | 0.004834941 | G6PD, GDF15, ZBTB16, PDGFB, CRYL1, HMOX1, CDH13, MVD, TPD52L1, PTGS2, PDLIM4, ZDHHC2 | |
| collagen binding | 9.152777778 | 0.009337216 | LUM, PDGFB, DCN, THBS4 | |
| chemokine activity | 13.35810811 | 0.020808064 | CXCL9, CCL21, CXCL1 | |
| benzaldehyde dehydrogenase (NAD+) activity | 82.375 | 0.023840283 | ALDH1A1, ALDH1A7 | |
| palmitoyl-CoA 9-desaturase activity | 82.375 | 0.023840283 | SCD, SCD2 | |
| heme binding | 4.476902174 | 0.025071838 | SRC, CYP1B1, HMOX1, CYP4A2, PTGS2 | |
| stearoyl-CoA 9-desaturase activity | 65.9 | 0.029711838 | SCD, SCD2 | |
| protein binding | 1.694571429 | 0.03195988 | CDKN1C, UNC5B, SRC, ZBTB16, PTGS2, PPP2CA, EFEMP1, CASP12, TRPV4, FASN, ADORA1, HMOX1, TSPO, CD9, CCDC141, PDLIM4, MPDZ, GRIA3 | |
| 3-chloroallyl aldehyde dehydrogenase activity | 54.91666667 | 0.035548404 | ALDH1A1, ALDH1A7 | |
| glyceraldehyde-3-phosphate dehydrogenase (NAD+) (non-phosphorylating) activity | 47.07142857 | 0.041350187 | ALDH1A1, ALDH1A7 | |
| KEGG | AMPK signaling pathway | 5.919354839 | 0.009421368 | PPP2CA, SCD, FASN, SCD2, ACACB |
| Alcoholic liver disease | 5.48088411 | 0.012249191 | SCD, FASN, SCD2, CXCL1, ACACB | |
| Metabolic pathways | 1.749653733 | 0.014414787 | MT-ND6, G6PD, CYP4A2, PTGS2, ACACB, LSS, PLA2G7, AACS, ACLY, SCD, FASN, ALDH1A1, ME1, SCD2, PNPLA3, CRYL1, HMOX1, MVD, ALDH1A7 | |
| PPAR signaling pathway | 6.882970743 | 0.019366476 | SCD, SCD2, ME1, CYP4A2 | |
| Cytokine-cytokine receptor interaction | 3.276395667 | 0.03364291 | CXCL9, TNFRSF12A, CCL21, GDF15, CXCL1, IL17RB | |
| Reactome | Metabolism of lipids | 2.817831233 | 0.004103554 | ACLY, SCD, FASN, PNPLA3, CYP1B1, TSPO, PNPLA5, MVD, PTGS2, LSS, AACS |
| G alpha (i) signalling events | 3.685106717 | 0.010239663 | PPP2CA, CXCL9, CCL21, SRC, ADORA1, CXCL1, TAS1R1 | |
| GPCR downstream signalling | 2.649684044 | 0.016729272 | PPP2CA, CXCL9, CCL21, SRC, ARHGEF16, ADORA1, CXCL1, TAS1R1, RHOC | |
| Metabolism | 1.670611517 | 0.019928616 | MT-ND6, G6PD, LUM, PTGS2, LSS, AACS, DCN, ACLY, SCD, FASN, ALDH1A1, PNPLA3, CRYL1, HMOX1, TSPO, CYP1B1, PNPLA5, MVD, SULT4A1 | |
| Fatty acyl-CoA biosynthesis | 12.85249042 | 0.021695752 | ACLY, SCD, FASN | |
| Fatty acid metabolism | 4.501476114 | 0.023057567 | ACLY, SCD, FASN, CYP1B1, PTGS2 | |
| G1 Phase | 9.808479532 | 0.035914728 | PPP2CA, CDKN1C, SRC | |
| Cyclin D associated events in G1 | 9.808479532 | 0.035914728 | PPP2CA, CDKN1C, SRC | |
| Chemokine receptors bind chemokines | 8.470959596 | 0.046907572 | CXCL9, CCL21, CXCL1 | |
| Signaling by GPCR | 2.162798195 | 0.048288983 | PPP2CA, CXCL9, CCL21, SRC, ARHGEF16, ADORA1, CXCL1, TAS1R1, RHOC |
Examination of lipogenesis and fatty acid oxidation genes revealed that lipogenic genes (Acly, Scd, Fasn) increased with aging and decreased with CR, while fatty acid oxidation genes showed opposite trends (Table 2). qRT-PCR was validated that PPARs and fatty acid oxidation genes (Ppara, Ppard, Cpt2, Cyp4a1, Acaa1a) were downregulated during aging but upregulated with CR, whereas the lipogenic genes (Acaca, Fasn) showed the opposite results (Supplementary Table 4 and Figure 4).
Table 2. Changes in the expression of genes related to lipid metabolism from RNA-Seq data in the Old vs. Young and Old-CR vs. Old datasets.
| Category | Genez | Old vs. Young | p-value | Old-CR vs. Old | p-value |
| Lipogenesis | Aacs | 3.972369982 | 3.30E-08 | −2.02791896 | 2.00E-03 |
| Acly | 1.777685362 | 9.50E-03 | −1.729074463 | 1.10E-02 | |
| Acaca | 1.945309895 | 4.90E-03 | 1.049716684 | 8.30E-01 | |
| Acacb | 1.558329159 | 4.90E-02 | −1.905275996 | 3.00E-03 | |
| Fasn | 2.203810232 | 4.20E-04 | −2.056227653 | 8.90E-04 | |
| Pnpla3 | 6.105036836 | 4.30E-14 | −2.887858391 | 1.80E-06 | |
| Pparg | −1.197478705 | 6.00E-01 | −1.148698355 | 7.40E-01 | |
| Scd | 1.777685362 | 9.80E-03 | −4.469148552 | 1.70E-11 | |
| Scd2 | 3.052518418 | 3.30E-05 | −2.099433367 | 2.90E-03 | |
| Fatty acid oxidation | Acox1 | −1.125058485 | 5.90E-01 | 1.494849249 | 6.30E-02 |
| Cpt1a | −1.021012126 | 9.20E-01 | 1.433955248 | 9.20E-02 | |
| Cpt2 | 1.205807828 | 4.00E-01 | 1.283425898 | 2.50E-01 | |
| Acadvl | −1.079228237 | 7.20E-01 | 1.042465761 | 8.50E-01 | |
| Hadhb | 1.222640278 | 3.70E-01 | 1.248330549 | 3.10E-01 | |
| Acadm | −1.172834949 | 4.70E-01 | −1.117287138 | 6.00E-01 | |
| Echs1 | −1.515716567 | 6.30E-02 | 1.292352831 | 2.40E-01 | |
| Hadh | −1.125058485 | 6.00E-01 | −1.189207115 | 4.30E-01 | |
| Ppara | −1.164733586 | 5.70E-01 | 1.394743666 | 1.70E-01 | |
| Ppard | −2.531513188 | 2.40E-04 | 1.310393404 | 2.90E-01 | |
| Acox1 | −1.125058485 | 5.90E-01 | 1.494849249 | 6.30E-02 |
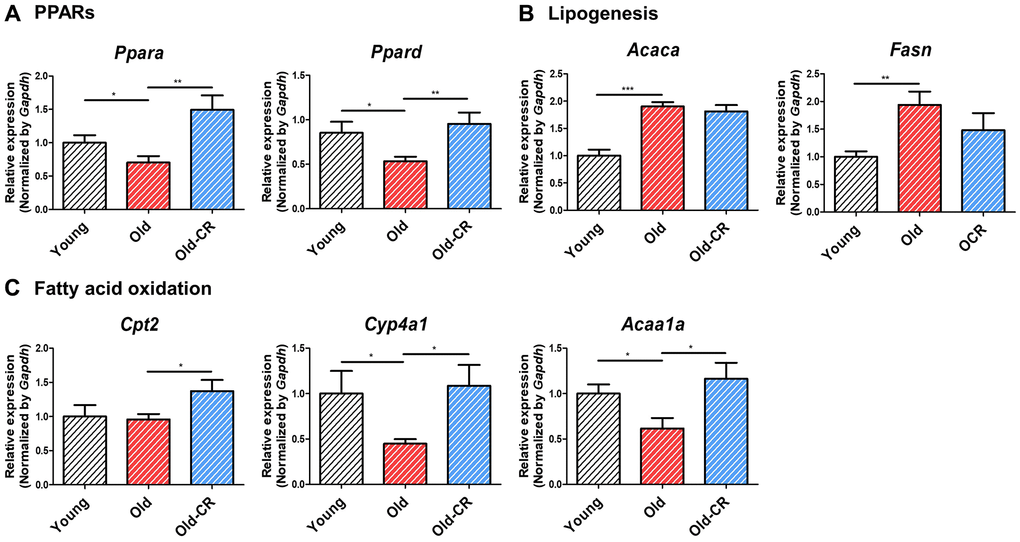
Figure 4. Relative mRNA expression of genes related to lipid metabolism that are differentially expressed during aging and CR (n = 6 per group). (A) Ppara and Ppard are downregulated in aging but upregulated in CR, (B) while the lipogenic genes Acaca and Fasn display opposite trends. (C) Fatty acid oxidation-related genes, such as Cpt2, Cyp4a1, and Acca1a, are downregulated in aging but upregulated in CR. Data are presented as mean ± SEM. *p < 0.05, **p < 0.01, and ***p < 0.001 between two groups.
Triglyceride (TG) levels were measured to assess the aging and CR effects on lipid metabolism. TG levels were significantly elevated in the Old group compared to Young (106.8 ± 11.23 vs. 82.28 ± 7.111 mg/dL; p < 0.05); but significantly reduced in the Old-CR compared to Old (40.73 ± 3.689 mg/dL; p < 0.0001) (Figure 5). These findings indicate that lipid synthesis genes increased with aging and decreased with CR, affecting lipid accumulation and suggesting that changes in lipogenesis and fatty acid oxidation are crucial in hepatic aging, with CR potentially counteracting these alterations.
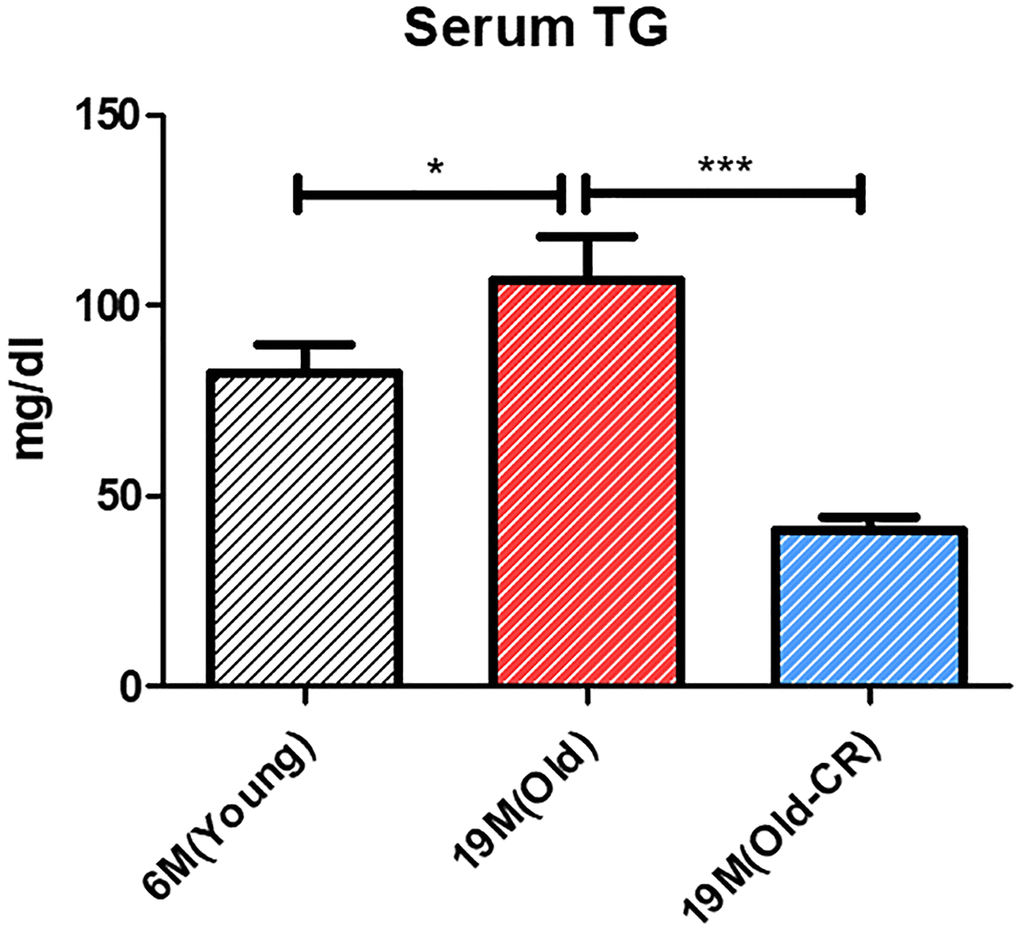
Figure 5. Effect of aging and CR on serum TG (n = 6 per group). Serum TG levels are increased in Old rats compared to those in Young rats and are decreased in Old-CR rats compared to those in Old rats (Young: 82.28 ± 7.111 mg/dL, Old: 106.8 ± 11.23 mg/dL, and Old-CR: 40.73 ± 3.689 mg/dL). Data are presented as mean ± SEM. Old-CR, old group under CR; TG, triglycerides.
Expression changes of the circadian gene Nr1d1 predicted as a key gene linking circadian rhythms and metabolism by CR during aging
Following lipid metabolism analysis, we examined circadian rhythm genes to identify aging- and CR-related expression changes (Table 3). Most circadian genes showed significant alterations in both Old vs. Young and Old-CR vs. Old datasets, with Cry1, Cry2, Nfil3, Nr1d1, and Rorc displaying opposite trends in both datasets. We focused on Nr1d1, a circadian repressor that increased with age. Furthermore, qRT-PCR validated circadian gene changes (Figure 6 and Supplementary Table 4), showing that Nr1d1 and Nr1d2 increased in the Old group compared to Young and decreased in the Old-CR compared to Old. Conversely, Bmal1 decreased with aging but increased with CR. Clock also decreased with aging. Other genes such as Rora, Rorc, Per1, Per2, Cry1, and Cry2 showed with aging and significant increases with CR.
Table 3. Changes in the expression of circadian genes from RNA-Seq data in the Old vs. Young and Old-CR vs. Old datasets.
| Old vs. Young | p-value | Old-CR vs. Old | p-value | |
| Bmal1 (Arntl) | −3.116658319 | 4.20E-06 | −9.51365692 | 9.20E-12 |
| Clock | −1.404444876 | 1.30E-01 | −1.879045498 | 5.40E-03 |
| Cry1 | −1.777685362 | 4.80E-02 | 4.75682846 | 4.90E-09 |
| Cry2 | −1.021012126 | 9.30E-01 | 2.099433367 | 1.00E-03 |
| Dbp | 12.55334557 | 2.60E-23 | 1.945309895 | 2.10E-03 |
| Nfil3 | −1.647182035 | 3.10E-02 | 1.569168196 | 4.70E-02 |
| Npas2 | −2.848100391 | 1.20E-05 | −23.58830748 | 2.50E-19 |
| Nr1d1 | 2.514026749 | 5.30E-05 | −1.214194884 | 3.80E-01 |
| Nr1d2 | 1.591072968 | 4.00E-02 | 1.484523571 | 7.00E-02 |
| Per1 | 1.021012126 | 9.20E-01 | 2.281527432 | 2.20E-04 |
| Per2 | 1.753211443 | 1.60E-02 | 4.228072162 | 2.00E-10 |
| Rora | −1.292352831 | 6.70E-01 | 1.2397077 | 7.80E-01 |
| Rorb | 1 | 1.00E+00 | 1 | 1.00E+00 |
| Rorc | −1.01395948 | 9.60E-01 | 2.0139111 | 1.50E-03 |
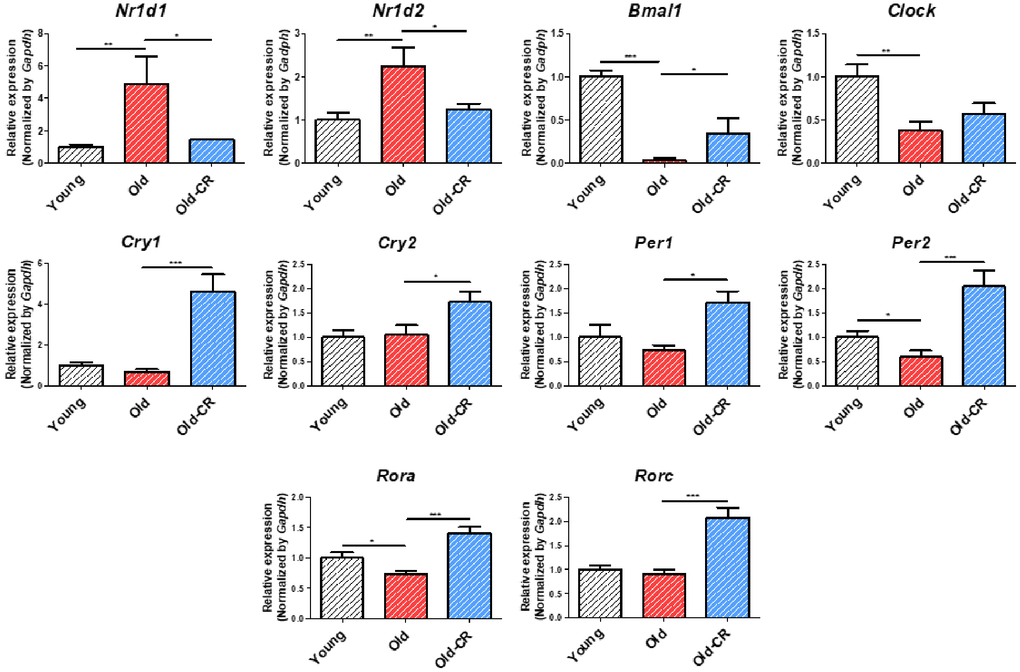
Figure 6. Relative mRNA expression of circadian genes that are differentially expressed during aging and CR (n = 6 per group). Nr1d1 and Nr1d2 are upregulated in aging but downregulated in CR, while Bmal1, Per2, and Rora display opposite trends. Clock is downregulated in aging. Cry1, Cry2, Per1, and Rorc are upregulated in CR. Data are presented as mean ± SEM. *p < 0.05, **p < 0.01, and ***p < 0.001 between two groups.
We constructed a protein-protein interaction (PPI) network based on circadian-associated genes, lipid metabolism-related genes, and aging-altered DEGs, trimmed to focus on Nr1d1 (Supplementary Figure 1 and Figure 7, Supplementary Table 5). Results confirmed that circadian rhythm and lipid metabolism genes were interconnected around Nr1d1, with Ppara among these connections. These analyses suggest that increased Nr1d1 expression during aging affects both circadian rhythm and lipid metabolism, which was ameliorated by CR.
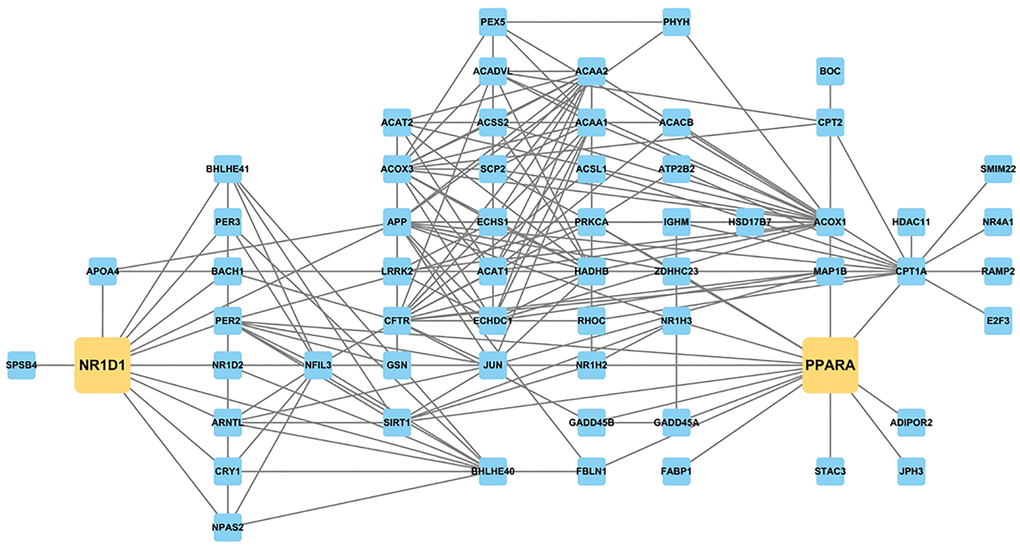
Figure 7. PPI network constructed with genes related to circadian rhythm and lipid metabolism and aging-altered DEGs, focusing on Nr1d1 and Ppara. NR1D1 is highly related to several circadian and metabolic regulators, such as Bmal1 (Arntl), Nr1d2, Cry1, Acox1, Cpt2, and Ppara.
Co-regulation of Nr1d1- and Pparα-induced age-related changes in lipid metabolism and its modulation by CR
To identify the genes closely associated with Nr1d1, we applied the RWR algorithm using Nr1d1 as the seed node (Supplementary Figure 1 and Supplementary Table 6). Among the top 100 genes, several circadian and lipid metabolism genes were identified, such as Cry1, Arntl, Nr1d2, Fasn, Pparg, and Acaca. Ppara ranked among top genes, indicating that Nr1d1 is closely associated with Pparα, a major metabolic regulator.
Western blotting detected Nr1d1 and Pparα levels (Figure 8). Nr1d1 protein levels were higher in the Old group than Young control but significantly downregulated in the Old-CR compared to Old. Conversely, Pparα protein levels were lower in Old than Young but increased with CR treatment. Immunohistochemistry assessed the Nr1d1 expression and localization in rat liver tissues (Figure 9). The Old group showed greater Nr1d1 intensity than Young, while intensity was significantly diminished in Old-CR compared to Old. These results suggest that the Nr1d1 and Pparα protein expression are affected by aging and CR with opposing trends, highlighting their potential as aging modulators.
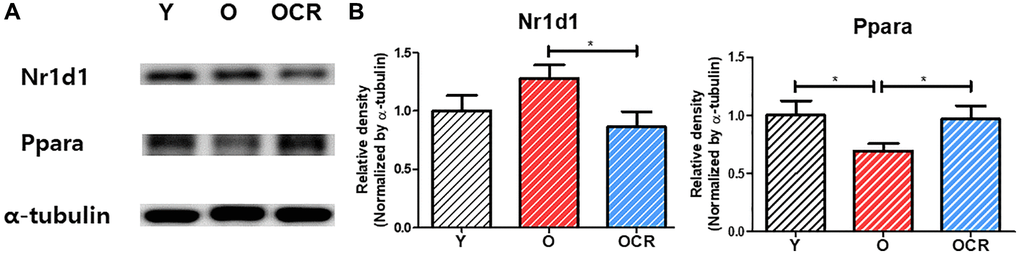
Figure 8. Protein expression of NR1D1 and PPARα during aging and CR (n = 6 per group). (A) Representative images of NR1D1 and PPARα bands. (B) Relative expression of NR1D1 and PPARα. The expression of NR1D1 increases during aging and decreases under CR. In contrast, the expression of PPARα is downregulated during aging but upregulated by CR. Data are presented as mean ± SEM. Abbreviations: Y: Young; O: Old; OCR: Old-CR. *p < 0.05 between two groups.
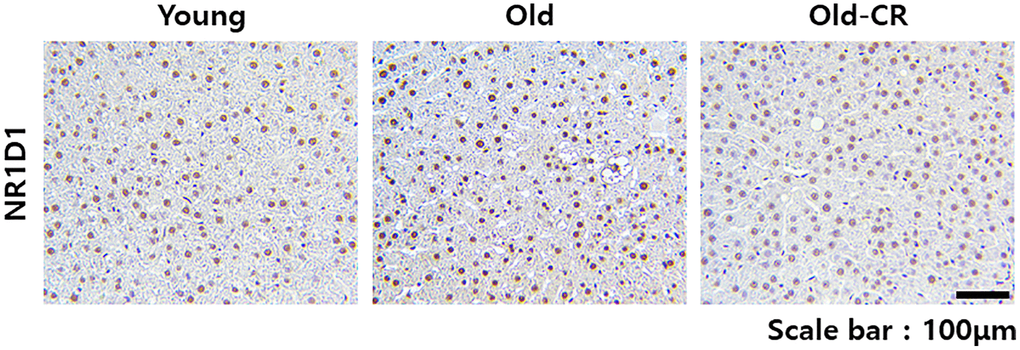
Figure 9. Immunohistochemical analysis of Nr1d1 expression in rat liver tissues during aging and CR. Nr1d1 protein expression is elevated in liver sections from aged mice compared to young controls, while CR intervention significantly attenuated this age-associated increase. Representative images of Nr1d1 staining of liver sections from the Young (left), Old (center), and Old-CR groups (right). Scale bar = 100 μm.
Effect of NR1D1 overexpression on PPARα binding to peroxisome proliferator response elements (PPRE) and the expression of circadian- and lipid metabolism-related genes
To investigate the relationship between NR1D1 and PPARα, we measured PPARα binding to PPRE in HepG2 cells using luciferase assay (Figure 10A). PPARα binding to PPRE decreased significantly in a dose-dependent manner with NR1D1 administration. To examine increased NR1D1 effects on gene expression during aging, we measured circadian rhythm and lipid metabolism gene expression in HepG2 cells overexpressing NR1D1 (Figure 10B). Results showed decreased expression of BMAL1, CLOCK, PER1, PER2, CRY1, and CRY2 in NR1D1-overexpressing cells and reduced expression of fatty acid oxidation genes (ACOX1, CPT2, PPARA). These results suggest that NR1D1 competitively binds to promoters, inhibiting PPARα binding to PPRE and reducing fatty acid oxidation gene expression.
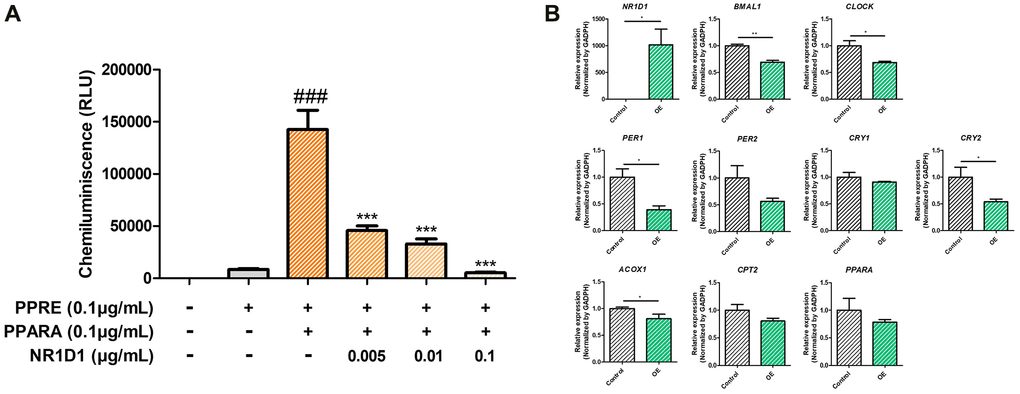
Figure 10. Effects of NR1D1 overexpression on (A) the PPRE binding activity of PPARα and (B) expression changes of circadian and lipid metabolic genes (n = 6 per group). HepG2 cells were transfected with pcDNA or the NR1D1 plasmid. The luciferase reporter assay measures the binding of PPARα to the PPRE element with varying amounts of NR1D1 expression vectors. PPRE binding of PPARα decreases in a NR1D1 dose-dependent manner. Furthermore, the circadian gene BMAL1, CLOCK, PER1/2, and CRY1/2 and lipid metabolic genes ACOX1, CPT2, and PPARA are decreased with Nr1d1 overexpression. Data are presented as mean ± SEM. ###, ***p < 0.001 between two groups. OE; overexpression.
Discussion
In this study, we identified key regulatory genes in the liver during aging and CR through transcriptomic and biological analyses. Transcriptomic analysis revealed changes in metabolic and circadian-related gene expression during aging and CR, with lipid metabolism showing opposite expression patterns. Notable changes in circadian gene expression were confirmed, particularly Nr1d1, which increased during aging and decreased with CR. PPI analysis predicted that Nr1d1 association with lipid metabolism genes, including Ppara. We demonstrated that NR1D1 competitively binds to the PPRE against PPARα in human cells, reducing fatty acid oxidation gene transcription. These findings suggest that Nr1d1 overexpression during aging interferes with Pparα, leading to hepatic lipid accumulation, which was alleviated by CR.
Our study confirmed significant changes in genes associated with lipid metabolism and circadian rhythm during aging and CR. These findings align with prior literature demonstrating that circadian rhythms and lipid metabolism undergo substantial alterations during hepatic aging, and directly linked to age-related metabolic diseases [29]. The complex interplay between metabolic, hormonal, and cellular mechanisms contributing to age-associated lipid accumulation has been well-documented [43, 44]. Aging modifies the circadian transcriptome of the liver, resulting in tissue-specific alterations in circadian gene expression patterns [25, 45].
CR, a well-known anti-aging strategy, positively affects circadian rhythm gene expression, helping synchronize circadian rhythms and protect against desynchronization that negatively affects longevity [46]. CR also enhances protein modifications such as acetylation, suggesting a pathway to slow aging by modulating liver metabolism [25]. These findings are consistent with our data indicating significant alterations in circadian rhythm and metabolism-related gene expression during hepatic aging and CR.
We confirmed that Nr1d1 is upregulated during aging but downregulated by CR. Our previous study demonstrated Nr1d1 overexpression during hepatic aging [41]. Other studies reported that Nr1d1 expression is associated with aging and lipid metabolism. For example, reducing Nr1d1 levels in aged mouse heart cells promotes growth and reduces cell death, offering a strategy to mitigate heart aging [47]. CR changes transcriptomic levels of longevity and circadian-related genes, including Nr1d1, thereby mitigating circadian disruption [48–51].
Nr1d1 plays a complex, context-dependent role in lipid metabolism regulation. While Nr1d1 suppression leads to increased hepatic lipid accumulation, particularly with high dietary fat intake [52, 53], other studies reported that NR1D1 positively regulates lipogenic genes such as SREBP-1c in hepG2 cells [54]. These seemingly contradictory findings likely reflect the multifaceted nature of Nr1d1 function, which varies depending on metabolic state, tissue context, and experimental conditions. Our data revealed that in the aging context, Nr1d1 overexpression disrupts the balance of lipid homeostasis by interfering with Pparα-mediated fatty acid oxidation, leading to lipid accumulation. This is consistent with studies showing that circadian clock proteins regulated by Nr1d1, including Bmal1, Clock, and cryptochromes, significantly regulate ketogenesis by interacting with Pparα in mouse models [55].
Our findings suggest that during aging, elevated Nr1d1 expression shifts this balance toward lipid accumulation by competitively inhibiting Pparα function, while CR restores metabolic balance by normalizing Nr1d1 levels. Although previous research on Nr1d1 overexpression has been limited, our study highlights a novel role for Nr1d1 in significantly influencing lipid metabolism. This expands the understanding of Nr1d1 functions beyond its established roles, emphasizing the importance of maintaining Nr1d1 homeostasis as a potential strategy for regulating circadian rhythms and metabolism. Further research is warranted to explore these mechanisms in greater detail.
Our study confirmed that both Pparα expression and activity decline during liver aging but are restored by CR. PPARs play crucial roles in lipid metabolism regulation and prevention of obesity and diabetes, which can modulate aging [56–58]. Pparα, a member of the PPAR family, plays a protective role against age-related lesions in vital organs, including the liver, kidneys, and heart, thereby mitigating aging progression [59, 60]. These studies indicate that age-related PPARα decreases can aggravate the aging effects, highlighting its role as an aging modulator. On the other hand, CR leads to metabolic adaptations where the body reduces lipogenesis and increases lipolysis and ketogenesis by activating Pparα [9, 61]. Considering the critical role of Pparα as a metabolic regulator, comprehensive understanding of its targets and regulatory mechanisms could provide a robust foundation for addressing metabolic decline during aging.
Pparα is influenced by circadian genes and reciprocally regulates these genes while modulating lipid metabolism. Bmal1, in cooperation with Clock, activates Ppara expression by binding to E-box elements and acts as an upstream regulator of Ppara gene influencing various metabolic processes [62, 63]. Conversely, PPARα directly regulates core circadian rhythm components, such as BMAL1 and NR1D1, illustrating their significant role in metabolism and daily bodily cycles [64]. These findings support our study’s assertion of Pparα-Nr1d1 interaction during aging by demonstrating that Pparα functions as a regulator of circadian rhythm components, including Nr1d1.
We confirmed that NR1D1 competitively binds to the PPRE and inhibits the binding of PPARα to PPRE in hepG2 cells. NR1D1 modulates gene expression by antagonizing the binding of PPARα to the PPRE, illustrating a convergence of NR1D1 and PPAR signaling pathways in the transcriptional regulation affecting lipid metabolism, including β-oxidation [65–67]. In mouse models, Nr1d1 represses downstream circadian genes like Bmal1 and Clock, while Pparα regulates the circadian clock by modulating Bmal1 [63, 68]. Bmal1/Clock reciprocally transactivates PPAR target genes via PPRE, indicating bidirectional influence of circadian and metabolic regulators [69]. These studies correspond with our findings that Nr1d1 plays a crucial role in modulating lipid metabolism by regulating fatty acid oxidation-related gene expression, which are principal Pparα targets.
In conclusion, our findings reveal that the co-regulation of Nr1d1 and Pparα plays a significant role in modulating hepatic lipid accumulation during aging, while CR may protect against this process, offering new potential strategies to mitigate the age-related metabolic decline with liver aging (Figure 11).
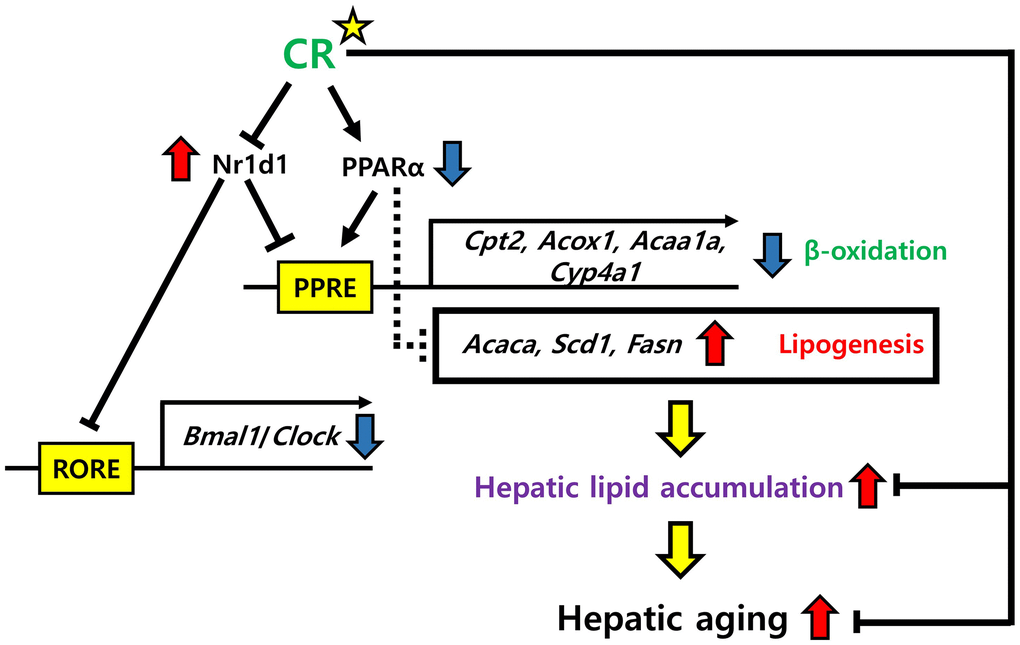
Figure 11. Possible mechanisms of NR1D1 and PPARα modulation of hepatic lipid metabolism in aging and CR. NR1D1 and PPARα competitively bind to PPRE. Upregulation of NR1D1 during aging represses the expression of genes involved in β-oxidation, such as Cpt2, Acox1, Acaa1a, and Cyp4a1 and further induces the expression of lipogenic genes, such as Acaca, Scd1, and Fasn. These transcriptomic changes lead to hepatic lipid accumulation, which aggravates hepatic aging. However, CR may protect against the deterioration of lipid metabolism in the liver by modulating NR1D1 and PPARα.
Materials and Methods
Animals
Young male Sprague Dawley (SD) rats (Young group; 6 months old), old male SD rats (Old group; 19 months old), and old male rats treated with caloric restriction (Old-CR group; 19 months old) were purchased from Samtako (Osan, Gyeonggi-do, Korea). The CR protocol followed established methodologies where old-CR rats received 60% of the average daily food intake measured after 1-week stabilization, maintained for 4 weeks. Young and old rats were provided ad libitum water and a normal chow diet (20% protein, 4.5% fat, 6% fibre, 7% ash, 0.5% calcium, 1% phosphorus) [14, 42]. Rats were kept at 23 ± 2°C, with 60 ± 5% humidity and a 12-h light/dark cycle. Tissues were frozen in liquid nitrogen for analysis. All experiments were approved by the Pusan National University Institutional Animal Care and Use Committee (approval number PNU-2015-1044).
RNA-Seq
Total RNA was extracted from liver samples using RiboEx reagent (GeneAll Biotechnology, Seoul, Korea). Equal quantities of RNA from each group (n = 5) were pooled for RNA-Seq analysis. cDNA libraries were prepared and sequenced using the MGI-T7 platform (MGI Tech Co., Shenzhen, China) with MGIEasy RNA Directional Library Prep Set. Adapter sequences were removed using Cutadapt (version 2.9) and quality filtered using Trimmomatic (version 0.39) [70, 71]. Reads shorter than 36 bp were excluded from analysis. Reads were aligned to the human reference genome (hg38) and Ensembl (version 102) using STAR (version 2.7.3a). STAR and RSEM (version 1.3.1) were used for alignment and quantification with default parameters [72, 73].
Differential expression analysis
Gene count data were normalized using the DESeq2 normalization methods. Differential expression analysis was performed using DESeq2 (version 1.30.1) [74]. Differentially expressed genes (DEGs) were identified using p-values < 0.05 and absolute fold changes ≥1.5 (Supplementary Table 1). Volcano plots were generated using VolcaNoseR web interface [75].
Gene ontology (GO) and pathway enrichment analyses
GO enrichment analysis examined biological functions, and KEGG pathway analysis investigated DEG-related signaling pathways. Both analyses were conducted using DAVID with statistical threshold of p < 0.05. Results are listed in Supplementary Table 2.
qRT-PCR
Primers were synthesized by Bioneer (Daejeon, Korea). Total RNA was isolated from rat liver tissues (20 mg) and NR1D1-overexpressing HepG2 cells using the RNeasy Mini Kit (Qiagen, Hilden, Germany) (n = 6 per group) and reverse-transcribed using cDNA synthesis kit (GenDEPOT, Baker, TX, USA). qRT-PCR was performed using SYBR Green (Bioneer, Daejeon, Korea) and CFX Connect System (Bio-Rad, Hercules, CA, USA). Unpaired Student’s t-test was used for two-group comparisons (NR1D1 overexpression vs. Control). When comparing multiple groups simultaneously, one-way analysis of variance (ANOVA) was performed followed by Tukey’s multiple comparison post-hoc test to control for family-wise error rate (Young vs. Old vs. Old-CR). Primer sequences are listed in Supplementary Table 4.
Serum biochemical analysis
Serum samples were prepared by centrifugation (4°C and 2,000×g for 15 min) after euthanasia. TG levels were measured using serum kits (Bioassay Systems, Hayward, CA, USA). Serum TG levels were compared between groups using one-way ANOVA followed by Tukey’s post-hoc test.
PPI network analysis
Mixed PPI database was constructed based on the interactions between five databases: HPRD, BioGRID, IntAct, MINT, and STRING [76–80]. Interactions with highest confidence (0.900) were selected from STRING, with all interactions extracted from the other databases. PPI network was constructed to identify hub molecules and examine DEG interactions. Cytoscape (version 3.10.1) visualized networks and calculated topological parameters including degree and betweenness centrality [81].
Random Walk with Restart (RWR) algorithm on the PPI network
RWR algorithm was applied for node ranking within the PPI network to identify pivotal genes near seed genes [82, 83]. The mathematical underpinning of RWR involves the utilization of a transition matrix (M) and probability vectors, where P signifies the vector denoting node probabilities. The updated equation is expressed, as follows:
Here, α represents the restart probability (damping factor), P(t) denotes the probability vector at iteration t, and Pinit is the initial probability vector. RWR computations used 0.8 restart probability with R package RandomWalkRestartMH [84]. Results are presented in Supplementary Table 6.
Western blotting
Total protein was extracted from rat liver tissues and boiled for 5 min in loading buffer containing 0.2% bromophenol blue, 125 mM Tris-HCl, 10% 2-mercaptoethanol, and 4% SDS (pH 6.8) (n = 6 per group). Equal amounts of protein (8–10 μg) were separated by SDS-PAGE using 10% gels and transferred to PVDF membranes at 25 V for 10 minutes using a semidry transfer. Membranes were blocked in 5% nonfat milk for 2 h, then immunoblotted overnight with primary antibodies (1:1,000) at 4°C. After washing, the membranes were incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies (1:10,000) for 1 h at room temperature. Protein detection was performed using enhanced chemiluminescence, and molecular weights were determined using a wide range of protein markers. Immunoblots were visualized using a chemiluminescent HRP substrate (Davinchchemi CAS-400) and analyzed using the ImageJ Software. The detailed procedure has been described previously [41]. Antibodies against NR1D1 (sc-100910), PPARα (sc-398394), and α-tubulin (sc-5286) were purchased from Santa Cruz (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Secondary antibodies (GTX213110-01 and GTX213111-01) were purchased from GeneTex (Irvine, CA, USA). Protein levels were compared between groups using one-way ANOVA followed by Tukey's post-hoc test.
Immunohistochemistry
Paraffin-embedded sections were deparaffinized of using xylene, followed by graded ethanol series (starting at 100% and ending at 70%) and rehydration. Antigen retrieval was performed using sodium citrate buffer (pH 6.0). Specimens were then incubated with Nr1d1 primary antibodies (Santa Cruz, sc-100910, 1:200) at 4°C overnight in a humidity-controlled environment. After PBS washing, sections were incubated with biotinylated secondary antibodies (1:10,000; VectorLabs) for 30 min at ambient temperature. Diaminobenzidine substrate with hematoxylin counterstaining was used for visualization. Microscopic examinations were performed using a Motic AE30/31 inverted microscope.
Cell culture
HepG2 cells were purchased from American Type Culture Collection and maintained in Dulbecco’s Modified Eagle Medium (Welgene; LM001-11) containing 10% fetal bovine serum (Gibco; S001-01), 100 U/mL penicillin, and 100 μg/mL streptomycin (Hyclone; SV30010) at 37°C in 5% CO2. Cells were cultured in sterile plastic plates (SPL, 20100).
Cell transfection for NR1D1 overexpression
To observe the change in mRNA expression with NR1D1 overexpression, HepG2 cells were transfected with pcDNA or the NR1D1 plasmid using Lipofectamine 3000 (Invitrogen; L3000015), according to the manufacturer’s instructions. The NR1D1 plasmid was kindly provided by Dr. Mi-Ock Lee (Seoul National University, Seoul, Republic of Korea).
Luciferase assay
To observe the binding of PPARα or NR1D1 to PPRE, HepG2 cells (1.5 × 104 cells/well) were grown in a 96-well plate in DMEM supplemented with 10% FBS. The PPRE-X3-TK-LUC plasmid (0.1 μg) purchased from BioCat company (Heidelberg, Land Baden-Württemberg, Germany), and PPARA and the NR1D1 plasmid were transfected using Lipofectamine 3000 (0.1 μL) and P3000 (0.2 μL) complexes in Opti-MEM, according to the manufacturer’s instructions. An empty pcDNA vector was added to ensure that equal amounts of plasmid DNA were used per transfection. Luciferase activity was measured using a ONE-Glo Luciferase Assay System (Promega, Madison, WI, USA) and a luminescence plate reader (Berthold Technologies GmbH & Co., Bad Wildbad, Germany). Data were analyzed using one-way ANOVA with Tukey’s post-hoc test for multiple comparisons.
Statistical analysis
Sample sizes (n = 6 per group for animal studies, n = 3 for cell culture experiments) were determined based on our previous studies to detect meaningful biological differences [27, 85]. All statistical analyses were performed using the GraphPad Prism software (version 5; GraphPad Software, La Jolla, CA, USA). Data are presented as mean ± standard error of the mean (SEM). Statistical significance was set at p < 0.05 for all analyses.
Supplementary Materials
Author Contributions
Noh, S.G. and Chung, H.Y. conceived and designed the experiments. Noh, S.G. analyzed the data and wrote the manuscript. Kim, H.W. and Kim, S. performed all in vitro and in vivo experiments. Noh, S.G. performed all in silico analyses. Yu, B.P., Yoon, J.H., Chung, K.W., Lee, J., and Chung, H.Y. critically revised the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Ethical Statement
Experimental work with animals was approved by the Pusan National University Institutional Animal Care and Use Committee (Approval Number: PNU-2015-1044) and was performed in compliance with guidelines on animal experimentation at our institution.
Funding
This work was supported by National Research Foundation grants funded by the Korean government (NRF-2023R1A2C2006035, RS-2023–00272618).
References
- 1. Khaltourina D, Matveyev Y, Alekseev A, Cortese F, Ioviţă A. Aging Fits the Disease Criteria of the International Classification of Diseases. Mech Ageing Dev. 2020; 189:111230. https://doi.org/10.1016/j.mad.2020.111230 [PubMed]
- 2. Anantharaju A, Feller A, Chedid A. Aging Liver. A review. Gerontology. 2002; 48:343–53. https://doi.org/10.1159/000065506 [PubMed]
- 3. Nagarajan SR, Paul-Heng M, Krycer JR, Fazakerley DJ, Sharland AF, Hoy AJ. Lipid and glucose metabolism in hepatocyte cell lines and primary mouse hepatocytes: a comprehensive resource for in vitro studies of hepatic metabolism. Am J Physiol Endocrinol Metab. 2019; 316:E578–89. https://doi.org/10.1152/ajpendo.00365.2018 [PubMed]
- 4. Sharabi K, Tavares CDJ, Puigserver P. Regulation of Hepatic Metabolism, Recent Advances, and Future Perspectives. Curr Diab Rep. 2019; 19:98. https://doi.org/10.1007/s11892-019-1224-4 [PubMed]
- 5. Radonjić T, Dukić M, Jovanović I, Zdravković M, Mandić O, Popadić V, Popović M, Nikolić N, Klašnja S, Divac A, Todorović Z, Branković M. Aging of Liver in Its Different Diseases. Int J Mol Sci. 2022; 23:13085. https://doi.org/10.3390/ijms232113085 [PubMed]
- 6. Morgan AE, Mc Auley MT. Cholesterol Homeostasis: An In Silico Investigation into How Aging Disrupts Its Key Hepatic Regulatory Mechanisms. Biology (Basel). 2020; 9:314. https://doi.org/10.3390/biology9100314 [PubMed]
- 7. Kim DH, Bang E, Jung HJ, Noh SG, Yu BP, Choi YJ, Chung HY. Anti-aging Effects of Calorie Restriction (CR) and CR Mimetics based on the Senoinflammation Concept. Nutrients. 2020; 12:422. https://doi.org/10.3390/nu12020422 [PubMed]
- 8. Flanagan EW, Most J, Mey JT, Redman LM. Calorie Restriction and Aging in Humans. Annu Rev Nutr. 2020; 40:105–33. https://doi.org/10.1146/annurev-nutr-122319-034601 [PubMed]
- 9. Kuhla A, Hahn S, Butschkau A, Lange S, Wree A, Vollmar B. Lifelong caloric restriction reprograms hepatic fat metabolism in mice. J Gerontol A Biol Sci Med Sci. 2014; 69:915–22. https://doi.org/10.1093/gerona/glt160 [PubMed]
- 10. Hu L, Xia X, Zong Y, Gu Y, Wei L, Yin J. Calorie Restriction Enhanced Glycogen Metabolism to Compensate for Lipid Insufficiency. Mol Nutr Food Res. 2022; 66:e2200182. https://doi.org/10.1002/mnfr.202200182 [PubMed]
- 11. Seo AY, Hofer T, Sung B, Judge S, Chung HY, Leeuwenburgh C. Hepatic oxidative stress during aging: effects of 8% long-term calorie restriction and lifelong exercise. Antioxid Redox Signal. 2006; 8:529–38. https://doi.org/10.1089/ars.2006.8.529 [PubMed]
- 12. Barzilai N, Banerjee S, Hawkins M, Chen W, Rossetti L. Caloric restriction reverses hepatic insulin resistance in aging rats by decreasing visceral fat. J Clin Invest. 1998; 101:1353–61. https://doi.org/10.1172/JCI485 [PubMed]
- 13. Kang MJ, Kim HJ, Kim HK, Lee JY, Kim DH, Jung KJ, Kim KW, Baik HS, Yoo MA, Yu BP, Chung HY. The effect of age and calorie restriction on HIF-1-responsive genes in aged liver. Biogerontology. 2005; 6:27–37. https://doi.org/10.1007/s10522-004-7381-z [PubMed]
- 14. Lee AK, Kim DH, Bang E, Choi YJ, Chung HY. β-Hydroxybutyrate Suppresses Lipid Accumulation in Aged Liver through GPR109A-mediated Signaling. Aging Dis. 2020; 11:777–90. https://doi.org/10.14336/AD.2019.0926 [PubMed]
- 15. Ogrodnik M, Miwa S, Tchkonia T, Tiniakos D, Wilson CL, Lahat A, Day CP, Burt A, Palmer A, Anstee QM, Grellscheid SN, Hoeijmakers JHJ, Barnhoorn S, et al. Cellular senescence drives age-dependent hepatic steatosis. Nat Commun. 2017; 8:15691. https://doi.org/10.1038/ncomms15691 [PubMed]
- 16. Panda S. Circadian physiology of metabolism. Science. 2016; 354:1008–15. https://doi.org/10.1126/science.aah4967 [PubMed]
- 17. Pietrocola F, Galluzzi L, Bravo-San Pedro JM, Madeo F, Kroemer G. Acetyl coenzyme A: a central metabolite and second messenger. Cell Metab. 2015; 21:805–21. https://doi.org/10.1016/j.cmet.2015.05.014 [PubMed]
- 18. Guan D, Lazar MA. Interconnections between circadian clocks and metabolism. J Clin Invest. 2021; 131:e148278. https://doi.org/10.1172/JCI148278 [PubMed]
- 19. Fang B, Everett LJ, Jager J, Briggs E, Armour SM, Feng D, Roy A, Gerhart-Hines Z, Sun Z, Lazar MA. Circadian enhancers coordinate multiple phases of rhythmic gene transcription in vivo. Cell. 2014; 159:1140–52. https://doi.org/10.1016/j.cell.2014.10.022 [PubMed]
- 20. Geronikolou SA, Pavlopoulou A, Uça Apaydin M, Albanopoulos K, Cokkinos DV, Chrousos G. Non-Hereditary Obesity Type Networks and New Drug Targets: An In Silico Approach. Int J Mol Sci. 2024; 25:7684. https://doi.org/10.3390/ijms25147684 [PubMed]
- 21. Tevy MF, Giebultowicz J, Pincus Z, Mazzoccoli G, Vinciguerra M. Aging signaling pathways and circadian clock-dependent metabolic derangements. Trends Endocrinol Metab. 2013; 24:229–37. https://doi.org/10.1016/j.tem.2012.12.002 [PubMed]
- 22. Kondratova AA, Kondratov RV. The circadian clock and pathology of the ageing brain. Nat Rev Neurosci. 2012; 13:325–35. https://doi.org/10.1038/nrn3208 [PubMed]
- 23. Froy O. Metabolism and circadian rhythms--implications for obesity. Endocr Rev. 2010; 31:1–24. https://doi.org/10.1210/er.2009-0014 [PubMed]
- 24. Xu W, Li X. Aging Disrupts Circadian Rhythms in Mouse Liver Mitochondria. Molecules. 2023; 28:4432. https://doi.org/10.3390/molecules28114432 [PubMed]
- 25. Sato S, Solanas G, Peixoto FO, Bee L, Symeonidi A, Schmidt MS, Brenner C, Masri S, Benitah SA, Sassone-Corsi P. Circadian Reprogramming in the Liver Identifies Metabolic Pathways of Aging. Cell. 2017; 170:664–77.e11. https://doi.org/10.1016/j.cell.2017.07.042 [PubMed]
- 26. Sadria M, Layton AT. Aging affects circadian clock and metabolism and modulates timing of medication. iScience. 2021; 24:102245. https://doi.org/10.1016/j.isci.2021.102245 [PubMed]
- 27. Noh SG, Jung HJ, Kim S, Arulkumar R, Chung KW, Park D, Choi YJ, Chung HY. Sex-Mediated Differences in TNF Signaling-and ECM-Related Gene Expression in Aged Rat Kidney. Biol Pharm Bull. 2023; 46:552–62. https://doi.org/10.1248/bpb.b22-00601 [PubMed]
- 28. Johnson AA, Stolzing A. The role of lipid metabolism in aging, lifespan regulation, and age-related disease. Aging Cell. 2019; 18:e13048. https://doi.org/10.1111/acel.13048 [PubMed]
- 29. Chung KW. Advances in Understanding of the Role of Lipid Metabolism in Aging. Cells. 2021; 10:880. https://doi.org/10.3390/cells10040880 [PubMed]
- 30. Song R, Hu M, Qin X, Qiu L, Wang P, Zhang X, Liu R, Wang X. The Roles of Lipid Metabolism in the Pathogenesis of Chronic Diseases in the Elderly. Nutrients. 2023; 15:3433. https://doi.org/10.3390/nu15153433 [PubMed]
- 31. An HJ, Lee B, Kim SM, Kim DH, Chung KW, Ha SG, Park KC, Park YJ, Kim SJ, Yun HY, Chun P, Yu BP, Moon HR, Chung HY. A PPAR Pan Agonist, MHY2013 Alleviates Age-Related Hepatic Lipid Accumulation by Promoting Fatty Acid Oxidation and Suppressing Inflammation. Biol Pharm Bull. 2018; 41:29–35. https://doi.org/10.1248/bpb.b17-00371 [PubMed]
- 32. Li X, Wang J, Wang L, Gao Y, Feng G, Li G, Zou J, Yu M, Li YF, Liu C, Yuan XW, Zhao L, Ouyang H, et al. Lipid metabolism dysfunction induced by age-dependent DNA methylation accelerates aging. Signal Transduct Target Ther. 2022; 7:162. https://doi.org/10.1038/s41392-022-00964-6 [PubMed]
- 33. Frith J, Day CP, Henderson E, Burt AD, Newton JL. Non-alcoholic fatty liver disease in older people. Gerontology. 2009; 55:607–13. https://doi.org/10.1159/000235677 [PubMed]
- 34. Sheedfar F, Sung MM, Aparicio-Vergara M, Kloosterhuis NJ, Miquilena-Colina ME, Vargas-Castrillón J, Febbraio M, Jacobs RL, de Bruin A, Vinciguerra M, García-Monzón C, Hofker MH, Dyck JR, Koonen DP. Increased hepatic CD36 expression with age is associated with enhanced susceptibility to nonalcoholic fatty liver disease. Aging (Albany NY). 2014; 6:281–95. https://doi.org/10.18632/aging.100652 [PubMed]
- 35. Raghothama C, Harsha HC, Prasad CK, Pandey A. Bioinformatics and proteomics approaches for aging research. Biogerontology. 2005; 6:227–32. https://doi.org/10.1007/s10522-005-2617-0 [PubMed]
- 36. Zierer J, Menni C, Kastenmüller G, Spector TD. Integration of 'omics' data in aging research: from biomarkers to systems biology. Aging Cell. 2015; 14:933–44. https://doi.org/10.1111/acel.12386 [PubMed]
- 37. Li R, Li Y, Tian M, Zhang H, Lou L, Liu K, Zhang J, Zhao Y, Zhang J, Le S, Fu X, Zhou Y, Li W, et al. Comparative proteomic profiling reveals a pathogenic role for the O-GlcNAcylated AIMP2-PARP1 complex in aging-related hepatic steatosis in mice. FEBS Lett. 2022; 596:128–45. https://doi.org/10.1002/1873-3468.14242 [PubMed]
- 38. Zhang Y, Xiang D, Hu X, Ruan Q, Wang L, Bao Z. Identification and study of differentially expressed miRNAs in aged NAFLD rats based on high-throughput sequencing. Ann Hepatol. 2020; 19:302–12. https://doi.org/10.1016/j.aohep.2019.12.003 [PubMed]
- 39. Hong SE, Heo HS, Kim DH, Kim MS, Kim CH, Lee J, Yoo MA, Yu BP, Leeuwenburgh C, Chung HY. Revealing system-level correlations between aging and calorie restriction using a mouse transcriptome. Age (Dordr). 2010; 32:15–30. https://doi.org/10.1007/s11357-009-9106-3 [PubMed]
- 40. Park D, Lee EK, Jang EJ, Jeong HO, Kim BC, Ha YM, Hong SE, Yu BP, Chung HY. Identification of the dichotomous role of age-related LCK in calorie restriction revealed by integrative analysis of cDNA microarray and interactome. Age (Dordr). 2013; 35:1045–60. https://doi.org/10.1007/s11357-012-9426-6 [PubMed]
- 41. Noh SG, Jung HJ, Kim S, Arulkumar R, Kim DH, Park D, Chung HY. Regulation of Circadian Genes Nr1d1 and Nr1d2 in Sex-Different Manners during Liver Aging. Int J Mol Sci. 2022; 23:10032. https://doi.org/10.3390/ijms231710032 [PubMed]
- 42. Kim CH, Lee EK, Choi YJ, An HJ, Jeong HO, Park D, Kim BC, Yu BP, Bhak J, Chung HY. Short-term calorie restriction ameliorates genomewide, age-related alterations in DNA methylation. Aging Cell. 2016; 15:1074–81. https://doi.org/10.1111/acel.12513 [PubMed]
- 43. Seo E, Kang H, Choi H, Choi W, Jun HS. Reactive oxygen species-induced changes in glucose and lipid metabolism contribute to the accumulation of cholesterol in the liver during aging. Aging Cell. 2019; 18:e12895. https://doi.org/10.1111/acel.12895 [PubMed]
- 44. Zhao L, Zou X, Feng Z, Luo C, Liu J, Li H, Chang L, Wang H, Li Y, Long J, Gao F, Liu J. Evidence for association of mitochondrial metabolism alteration with lipid accumulation in aging rats. Exp Gerontol. 2014; 56:3–12. https://doi.org/10.1016/j.exger.2014.02.001 [PubMed]
- 45. Solanas G, Peixoto FO, Perdiguero E, Jardí M, Ruiz-Bonilla V, Datta D, Symeonidi A, Castellanos A, Welz PS, Caballero JM, Sassone-Corsi P, Muñoz-Cánoves P, Benitah SA. Aged Stem Cells Reprogram Their Daily Rhythmic Functions to Adapt to Stress. Cell. 2017; 170:678–92.e20. https://doi.org/10.1016/j.cell.2017.07.035 [PubMed]
- 46. Derous D, Mitchell SE, Wang L, Green CL, Wang Y, Chen L, Han JJ, Promislow DEL, Lusseau D, Douglas A, Speakman JR. The effects of graded levels of calorie restriction: XI. Evaluation of the main hypotheses underpinning the life extension effects of CR using the hepatic transcriptome. Aging (Albany NY). 2017; 9:1770–824. https://doi.org/10.18632/aging.101269 [PubMed]
- 47. Pu S, Wang Q, Liu Q, Zhao H, Zhou Z, Wu Q. Nr1d1 Mediated Cell Senescence in Mouse Heart-Derived Sca-1+CD31- Cells. Int J Mol Sci. 2022; 23:12455. https://doi.org/10.3390/ijms232012455 [PubMed]
- 48. Makwana K, Gosai N, Poe A, Kondratov RV. Calorie restriction reprograms diurnal rhythms in protein translation to regulate metabolism. FASEB J. 2019; 33:4473–89. https://doi.org/10.1096/fj.201802167R [PubMed]
- 49. Chen R, Zhang Z, Ma J, Liu B, Huang Z, Hu G, Huang J, Xu Y, Wang GZ. Circadian-driven tissue specificity is constrained under caloric restricted feeding conditions. Commun Biol. 2024; 7:752. https://doi.org/10.1038/s42003-024-06421-0 [PubMed]
- 50. Patel SA, Velingkaar N, Makwana K, Chaudhari A, Kondratov R. Calorie restriction regulates circadian clock gene expression through BMAL1 dependent and independent mechanisms. Sci Rep. 2016; 6:25970. https://doi.org/10.1038/srep25970 [PubMed]
- 51. Katewa SD, Akagi K, Bose N, Rakshit K, Camarella T, Zheng X, Hall D, Davis S, Nelson CS, Brem RB, Ramanathan A, Sehgal A, Giebultowicz JM, Kapahi P. Peripheral Circadian Clocks Mediate Dietary Restriction-Dependent Changes in Lifespan and Fat Metabolism in Drosophila. Cell Metab. 2016; 23:143–54. https://doi.org/10.1016/j.cmet.2015.10.014 [PubMed]
- 52. Na H, Lee H, Lee MH, Lim HJ, Kim HJ, Jeon Y, Kang HL, Lee MO. Deletion of exons 3 and 4 in the mouse Nr1d1 gene worsens high-fat diet-induced hepatic steatosis. Life Sci. 2016; 166:13–9. https://doi.org/10.1016/j.lfs.2016.10.003 [PubMed]
- 53. Hunter AL, Pelekanou CE, Barron NJ, Northeast RC, Grudzien M, Adamson AD, Downton P, Cornfield T, Cunningham PS, Billaud JN, Hodson L, Loudon AS, Unwin RD, et al. Adipocyte NR1D1 dictates adipose tissue expansion during obesity. Elife. 2021; 10:e63324. https://doi.org/10.7554/eLife.63324 [PubMed]
- 54. Berthier A, Vinod M, Porez G, Steenackers A, Alexandre J, Yamakawa N, Gheeraert C, Ploton M, Maréchal X, Dubois-Chevalier J, Hovasse A, Schaeffer-Reiss C, Cianférani S, et al. Combinatorial regulation of hepatic cytoplasmic signaling and nuclear transcriptional events by the OGT/REV-ERBα complex. Proc Natl Acad Sci U S A. 2018; 115:E11033–42. https://doi.org/10.1073/pnas.1805397115 [PubMed]
- 55. Mezhnina V, Ebeigbe OP, Velingkaar N, Poe A, Sandlers Y, Kondratov RV. Circadian clock controls rhythms in ketogenesis by interfering with PPARα transcriptional network. Proc Natl Acad Sci U S A. 2022; 119:e2205755119. https://doi.org/10.1073/pnas.2205755119 [PubMed]
- 56. Chung JH, Seo AY, Chung SW, Kim MK, Leeuwenburgh C, Yu BP, Chung HY. Molecular mechanism of PPAR in the regulation of age-related inflammation. Ageing Res Rev. 2008; 7:126–36. https://doi.org/10.1016/j.arr.2008.01.001 [PubMed]
- 57. Masternak MM, Bartke A. PPARs in Calorie Restricted and Genetically Long-Lived Mice. PPAR Res. 2007; 2007:28436. https://doi.org/10.1155/2007/28436 [PubMed]
- 58. Nunn AV, Bell J, Barter P. The integration of lipid-sensing and anti-inflammatory effects: how the PPARs play a role in metabolic balance. Nucl Recept. 2007; 5:1. https://doi.org/10.1186/1478-1336-5-1 [PubMed]
- 59. Howroyd P, Swanson C, Dunn C, Cattley RC, Corton JC. Decreased longevity and enhancement of age-dependent lesions in mice lacking the nuclear receptor peroxisome proliferator-activated receptor alpha (PPARalpha). Toxicol Pathol. 2004; 32:591–9. https://doi.org/10.1080/01926230490515283 [PubMed]
- 60. Corton JC, Anderson SP, Stauber A. Central role of peroxisome proliferator-activated receptors in the actions of peroxisome proliferators. Annu Rev Pharmacol Toxicol. 2000; 40:491–518. https://doi.org/10.1146/annurev.pharmtox.40.1.491 [PubMed]
- 61. Shimokawa I, Higami Y. Leptin signaling and aging: insight from caloric restriction. Mech Ageing Dev. 2001; 122:1511–9. https://doi.org/10.1016/s0047-6374(01)00284-6 [PubMed]
- 62. Oishi K, Shirai H, Ishida N. CLOCK is involved in the circadian transactivation of peroxisome-proliferator-activated receptor alpha (PPARalpha) in mice. Biochem J. 2005; 386:575–81. https://doi.org/10.1042/BJ20041150 [PubMed]
- 63. Canaple L, Rambaud J, Dkhissi-Benyahya O, Rayet B, Tan NS, Michalik L, Delaunay F, Wahli W, Laudet V. Reciprocal regulation of brain and muscle Arnt-like protein 1 and peroxisome proliferator-activated receptor alpha defines a novel positive feedback loop in the rodent liver circadian clock. Mol Endocrinol. 2006; 20:1715–27. https://doi.org/10.1210/me.2006-0052 [PubMed]
- 64. Chen L, Yang G. PPARs Integrate the Mammalian Clock and Energy Metabolism. PPAR Res. 2014; 2014:653017. https://doi.org/10.1155/2014/653017 [PubMed]
- 65. Kassam A, Capone JP, Rachubinski RA. Orphan nuclear hormone receptor RevErbalpha modulates expression from the promoter of the hydratase-dehydrogenase gene by inhibiting peroxisome proliferator-activated receptor alpha-dependent transactivation. J Biol Chem. 1999; 274:22895–900. https://doi.org/10.1074/jbc.274.32.22895 [PubMed]
- 66. Harding HP, Lazar MA. The monomer-binding orphan receptor Rev-Erb represses transcription as a dimer on a novel direct repeat. Mol Cell Biol. 1995; 15:4791–802. https://doi.org/10.1128/MCB.15.9.4791 [PubMed]
- 67. Duez H, Staels B. Rev-erb alpha gives a time cue to metabolism. FEBS Lett. 2008; 582:19–25. https://doi.org/10.1016/j.febslet.2007.08.032 [PubMed]
- 68. Chatterjee S, Yin H, Li W, Lee J, Yechoor VK, Ma K. The Nuclear Receptor and Clock Repressor Rev-erbα Suppresses Myogenesis. Sci Rep. 2019; 9:4585. https://doi.org/10.1038/s41598-019-41059-7 [PubMed]
- 69. Inoue I, Shinoda Y, Ikeda M, Hayashi K, Kanazawa K, Nomura M, Matsunaga T, Xu H, Kawai S, Awata T, Komoda T, Katayama S. CLOCK/BMAL1 is involved in lipid metabolism via transactivation of the peroxisome proliferator-activated receptor (PPAR) response element. J Atheroscler Thromb. 2005; 12:169–74. https://doi.org/10.5551/jat.12.169 [PubMed]
- 70. Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011; 17:3. https://doi.org/10.14806/ej.17.1.200
- 71. Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014; 30:2114–20. https://doi.org/10.1093/bioinformatics/btu170 [PubMed]
- 72. Dobin A, Davis CA, Schlesinger F, Drenkow J, Zaleski C, Jha S, Batut P, Chaisson M, Gingeras TR. STAR: ultrafast universal RNA-seq aligner. Bioinformatics. 2013; 29:15–21. https://doi.org/10.1093/bioinformatics/bts635 [PubMed]
- 73. Li B, Dewey CN. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics. 2011; 12:323. https://doi.org/10.1186/1471-2105-12-323 [PubMed]
- 74. Robinson MD, Oshlack A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010; 11:R25. https://doi.org/10.1186/gb-2010-11-3-r25 [PubMed]
- 75. Goedhart J, Luijsterburg MS. VolcaNoseR is a web app for creating, exploring, labeling and sharing volcano plots. Sci Rep. 2020; 10:20560. https://doi.org/10.1038/s41598-020-76603-3 [PubMed]
- 76. Keshava Prasad TS, Goel R, Kandasamy K, Keerthikumar S, Kumar S, Mathivanan S, Telikicherla D, Raju R, Shafreen B, Venugopal A, Balakrishnan L, Marimuthu A, Banerjee S, et al. Human Protein Reference Database--2009 update. Nucleic Acids Res. 2009; 37:D767–72. https://doi.org/10.1093/nar/gkn892 [PubMed]
- 77. Oughtred R, Rust J, Chang C, Breitkreutz BJ, Stark C, Willems A, Boucher L, Leung G, Kolas N, Zhang F, Dolma S, Coulombe-Huntington J, Chatr-Aryamontri A, et al. The BioGRID database: A comprehensive biomedical resource of curated protein, genetic, and chemical interactions. Protein Sci. 2021; 30:187–200. https://doi.org/10.1002/pro.3978 [PubMed]
- 78. Del Toro N, Shrivastava A, Ragueneau E, Meldal B, Combe C, Barrera E, Perfetto L, How K, Ratan P, Shirodkar G, Lu O, Mészáros B, Watkins X, et al. The IntAct database: efficient access to fine-grained molecular interaction data. Nucleic Acids Res. 2022; 50:D648–53. https://doi.org/10.1093/nar/gkab1006 [PubMed]
- 79. Licata L, Briganti L, Peluso D, Perfetto L, Iannuccelli M, Galeota E, Sacco F, Palma A, Nardozza AP, Santonico E, Castagnoli L, Cesareni G. MINT, the molecular interaction database: 2012 update. Nucleic Acids Res. 2012; 40:D857–61. https://doi.org/10.1093/nar/gkr930 [PubMed]
- 80. Szklarczyk D, Kirsch R, Koutrouli M, Nastou K, Mehryary F, Hachilif R, Gable AL, Fang T, Doncheva NT, Pyysalo S, Bork P, Jensen LJ, von Mering C. The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023; 51:D638–46. https://doi.org/10.1093/nar/gkac1000 [PubMed]
- 81. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003; 13:2498–504. https://doi.org/10.1101/gr.1239303 [PubMed]
- 82. Köhler S, Bauer S, Horn D, Robinson PN. Walking the interactome for prioritization of candidate disease genes. Am J Hum Genet. 2008; 82:949–58. https://doi.org/10.1016/j.ajhg.2008.02.013 [PubMed]
- 83. Zhu L, Su F, Xu Y, Zou Q. Network-based method for mining novel HPV infection related genes using random walk with restart algorithm. Biochim Biophys Acta Mol Basis Dis. 2018; 1864:2376–83. https://doi.org/10.1016/j.bbadis.2017.11.021 [PubMed]
- 84. Valdeolivas A, Tichit L, Navarro C, Perrin S, Odelin G, Levy N, Cau P, Remy E, Baudot A. Random walk with restart on multiplex and heterogeneous biological networks. Bioinformatics. 2019; 35:497–505. https://doi.org/10.1093/bioinformatics/bty637 [PubMed]
- 85. Ha S, Kim HW, Kim KM, Kim BM, Kim J, Son M, Kim D, Kim MJ, Yoo J, Yu HS, Jung YS, Lee J, Chung HY, Chung KW. PAR2-mediated cellular senescence promotes inflammation and fibrosis in aging and chronic kidney disease. Aging Cell. 2024; 23:e14184. https://doi.org/10.1111/acel.14184 [PubMed]